


-
胶质瘤是人类中最常见的恶性原发性颅内癌,尽管目前有多种治疗模式,仍然是人类恶性肿瘤中致死率较高的肿瘤之一[1]。胶质瘤具有独特的特征,包括肿瘤持续进展和强侵袭性及血管生成。其中,血管生成被认为是导致胶质瘤分级和恶性增加的原因[2]。另外, 胶质瘤还有无包膜浸润性生长特性,肿瘤的级别越高其浸润力越强,并且与肿瘤血管化相关[3]。肿瘤血管对肿瘤细胞的生长、增殖及运动过程中起着至关重要的作用。但是,在胶质瘤中,肿瘤血管对胶质瘤细胞的运动是否有促进作用,以及其中的分子机制并不是十分明确,因此,本实验通过体外分离培养胶质瘤血管内皮细胞,将其与胶质瘤细胞共培养,初步探讨肿瘤血管内皮细胞(GVEC)对胶质瘤细胞迁移能力影响及其作用机制。
-
本实验标本均取自2015-2016年我科手术治疗标本,病理诊断均由病理科根据WHO神经系统肿瘤分类作出,共收集10例WHO分级Ⅳ级胶质母细胞瘤标本。所有的标本去除表面的残留血液,放入含青、链霉素的无血清培养基中,4 ℃保存,6 h内分离内皮细胞培养。
-
CHG5细胞系购自中科院细胞库;RPMI-1640细胞培养基购自Invitrogen公司;无支原体新生胎牛血清为Hyclone生物工程公司产品;胰酶购自Thermo公司;RAC1、Cdc42、β-actin抗体为Santa Curz公司产品;细胞骨架免疫荧光试剂盒购自武汉博士德生物公司。
-
用PBS和抗生素洗涤肿瘤组织,并用手术刀切碎。然后将样品与0.625 Wu/mL释放酶在37 ℃下孵育1~2 h,并在无菌D-PBS中稀释后,悬浮液离心(276 g,10 min), 室温下将上清液分离。然后将细胞接种到胶原蛋白包被的烧瓶上,并补充内皮增殖介质(endothelial proliferation medium, EndoPM)放入5% CO2、1% O2、37 ℃环境中培养[4]。
-
将筛选出的内皮细胞和胶质瘤细胞系CHG5细胞培养于含10 %胎牛血清的RPMI-1640完全培养基中,置于37 ℃、5% CO2的培养箱内。细胞经传代和收集,取对数生长期细胞进行实验。实验分3组,CHG5单独培养(CHG5组)、正常血管内皮细胞(VEC)与CHG5共培养(VEC+CHG5组)和GVEC与CHG5共培养(GVEC+CHG5组)。
-
将分类的血管内皮细胞与胶质瘤细胞共培养于带Transwell微孔膜的24孔培养板中。Transwell将培养板分为上下两个小室,将肿瘤内皮细胞置于Transwell下室,CHG5细胞置于上室,加入500 μL含有10% FBS条件培养基,37 ℃培养箱,孵育24 h;培养24 h后,用PBS冲洗Transwell 2遍,去除上表面细胞,用4%多聚甲醛固定细胞,室温15 min;迁移至滤膜下面的细胞用结晶紫(0.1%)染色后PBS洗2遍,400倍显微镜下观察,随机计数10个视野内的细胞数,根据迁移至滤膜下表面的细胞数目的多少反映肿瘤细胞迁移能力的高低。计算平均值,进行统计学分析。
-
用上述方法共培养血管内皮细胞与胶质瘤细胞,为了便于细胞骨架荧光染色,此实验将胶质瘤细胞置于Transwell培养板下室,细胞在盖片上生长融合到95%~100%时,从培养箱中取出。用预温的PBS洗3次,-20 ℃甲醇固定2~3 min,再用PBS洗3次,每次3 min,5%BSA室温封闭10 min,加一抗(用0.1% BSA的PBS稀释,稀释比例为1:200)放在湿盒里,4 ℃过夜。PBS洗3次,每次2 min。加生物素化二抗(用0.1% BSA的PBS稀释,稀释比例为1:100),37 ℃孵育30 min。PBS洗3次,每次2 min。滴加SABC-Cy3(用0.1%BSA的PBS稀释,稀释比例为1:100),37 ℃孵育30 min,PBS洗4次,每次5 min,荧光显微镜观察照相。
-
与VEC、GVEC共培养后,收集CHG5细胞,用预冷PBS洗3次,加入适量的细胞裂解缓冲液,使细胞充分裂解,离心后收集蛋白。蛋白定量用紫外分光法测定。取等量蛋白,上样于SDS聚丙烯酞胺凝胶中,恒压电泳。电转法将凝胶上蛋白转移至PVDF膜。室温封闭(含5%脱脂奶粉的TBST)2 h。封闭后加入一抗(RAC1、Cdc42 1:1 000,β-actin 1:1 000),室温孵育2 h或4 ℃过夜,HRP偶联的二抗(1:1000)室温1 h,ECL显色。
-
采用方差分析和q检验。
-
用上述实验方法分离胶质瘤血管内皮细胞, 将细胞接种于培养瓶中, 经培养后, 倒置显微镜观察,内皮细胞生长良好, 细胞贴壁生长,形态完整, 折光性强(见图 1)。

图 1 胶质瘤血管内皮细胞分离培养结果
-
将GVEC、VEC分别与CHG5细胞通过Transwell小室共培养,进行体外细胞迁移实验,将迁移至Traswell微孔滤膜下的细胞,结晶紫染色,随机选取10个视野,计数平均值,实验重复3次,结果显示,与GVEC共培养的CHG5细胞的迁移能力显著高于与VEC共培养的CHG5细胞和单独培养的CHG5细胞(P<0.05)。
-
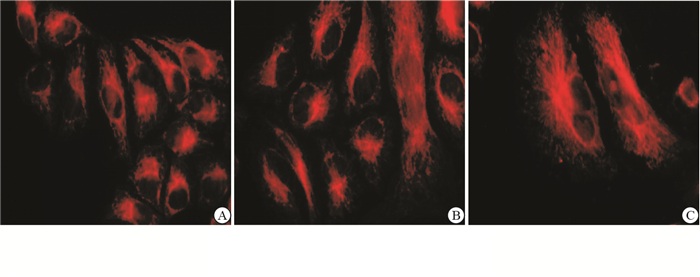
分别将与GVEC、VEC共培养的CHG5进行细胞骨架免疫荧光染色,结果显示,与GVEC共培养的CHG5细胞的微管蛋白在细胞核附近的微管蛋白较粗厚,呈圆环状,放射状排列;而与VEC共培养和单独培养的CHG5细胞微管蛋白弥漫分布呈细丝状(见图 2)。

图 2 CHG5细胞骨架免疫荧光染色结果
分组 迁移能力 F P MS组内 CHG5 57.33±2.08 VEC+CHG5 54.00±3.00 1342.28 < 0.01 6.542 GVEC+CHG5 149.33±2.51*# q检验:与CHG5组比较*P < 0.05;与VEC+CHG5组比较#P < 0.05 表 1 不同培养的胶质瘤CHG5细胞迁移能力的比较(x±s)
-
结果显示,与GVEC共培养的CHG5细胞Rac1和Cdc42的表达显著高于与VEC共培养的CHG5细胞和单独培养的CHG5细胞(P<0.05)(见表 2、图 3)。
分组 Rac1 Cdc42 CHG5 0.907±0.047 0.861±0.108 VEC+CHG5 0.977±0.011 1.113±0.013 GVEC+CHG5 1.156±0.021*# 1.444±0.065*# F 49.90 44.54 P < 0.01 < 0.01 MS组内 0.001 0.006 q检验:与CHG5组比较*P < 0.05;与VEC+CHG5组比较#P < 0.05 表 2 不同培养的胶质瘤CHG5细胞Rac1、Cdc42蛋白表达的比较(x±s)

图 3 CHG5细胞Rac1、Cdc42表达的情况
-
胶质母细胞瘤的一个独特特征是明显的血管生成,这有利于肿瘤生长和侵袭性[5]。肿瘤的血管在肿瘤发生发展的微环境中至关重要,除为肿瘤细胞提供营养外,还能增强肿瘤细胞的运动能力[6]。微管是动态细胞聚合物,对于许多细胞活动是必不可少的,包括有丝分裂、细胞分裂、信号转导、黏附、定向迁移、极化、囊泡和蛋白质递送至质膜和从质膜中回收,以及细胞形状和组织的重塑。细胞骨架,包括微管、肌动蛋白丝和中间丝(如波形蛋白、核纤层蛋白和角蛋白),不是一种稳定的结构,而是一种不断变化的细胞结构。虽然许多细胞能够通过单独的肌动球蛋白的作用产生细胞运动,但微管对于建立和维持极性,调控产生机械力和细胞黏附是非常重要的,因此,定向持续的细胞迁移和神经元寻路通常需要微管。微管细胞骨架本身是不对称组织的,以允许对细胞前部和后部的迁移机制进行差异调节。微管定位于细胞器如细胞核、中心体和高尔基体,并且与细胞的运动调控密切相关。本实验结果表明,与GVEC共培养的胶质瘤细胞胞核附近的微管蛋白较粗厚,呈圆环状,放射性排列;提示GVEC可诱导胶质瘤细胞的微管蛋白的重构、改变细胞的形态,促进细胞的迁移运动[7]。
Rho小GTP酶是单体20 000~30 000 GTP结合蛋白的Ras超家族的成员。这些蛋白质起分子开关的作用,调节各种细胞过程,如迁移、黏附和增殖。GDP结合的无活性形式和GTP结合的活性形式之间的循环受鸟嘌呤核苷酸交换因子、GTP酶活化蛋白和GDP-解离抑制剂的调节。在迄今鉴定的20种不同的哺乳动物Rho GTP酶中,Rac1和Cdc42已被最广泛地研究,已经证明,在各种类型的细胞中,Rac1和Cdc42蛋白质对细胞的迁移、黏附和增殖的调节起着关键的作用。细胞迁移过程中细胞的极化过程直接依赖于Rac1和Cdc42的局部活性。通常认为Rac1和Cdc42在迁移细胞的前沿起作用,其中它们诱导肌动蛋白突起。Rac1活化能诱导细胞边缘形成伪足,而Cdc42与丝状伪足的延伸相关,丝状伪足是从迁移细胞的前缘突出的肌动蛋白丝的细束和充当感知外部环境的探测器[8-9]。
在我们的实验中,GVEC通过促进胶质瘤细胞Rac1和Cdc42蛋白的表达,重调细胞内的微管蛋白,使细胞产生丝状伪足,促进细胞的迁移运动,提示肿瘤血管在胶质瘤细胞运动过程中起着至关重要的作用。目前,胶质瘤抗血管治疗已经取得良好的效果,本实验结果为抗血管治疗提供相关的理论基础,同时也为胶质瘤的治疗提供可能的分子靶向。
肿瘤血管内皮细胞对胶质瘤细胞迁移能力影响的机制研究
Study on the mechanism of the tumor vascular endothelial cells influencing the migration ability of glioma cells
-
摘要:
目的探讨肿瘤血管内皮细胞(GVEC)在胶质瘤肿瘤细胞迁移过程中的作用。 方法分别培养正常血管内皮细胞(VEC)及胶质瘤GVEC,在体外与胶质瘤细胞株(CGH5)共培养,并以单独培养的CHG5细胞作为对照。通过Transwell培养板迁移实验分析GVEC对CHG5细胞迁移能力的影响;免疫荧光染色观察GVEC对CHG5细胞骨架的影响;Western blotting检测共培养后CHG5细胞Rac1和Cdc42表达的变化。 结果CHG5与GVEC共培养后迁移能力显著高于与VEC共培养和单独培养的CHG5细胞(P < 0.05)。细胞骨架免疫荧光染色显示,与GVEC共培养的CHG5细胞的微管蛋白形态发生改变,细胞核附近微管蛋白较粗厚,呈圆环状,放射状排列。Western blotting结果表明,与GVEC共培养的CHG5细胞Rac1和Cdc42表达明显增高(P < 0.05),但与VEC共培养的CHG5细胞表达无明显变化(P>0.05)。 结论肿瘤细胞血管在胶质瘤细胞迁移的过程中至关重要,GVEC可以通过改变细胞微管蛋白结构,并上调Rac1和Cdc42蛋白的表达,促进胶质瘤细胞的迁移运动。 Abstract:ObjectiveTo investigate the role of tumor vascular endothelial cells (GVEC) in the migration of glioma tumor cells. MethodsThe vascular endothelial cells (VEC) and GVEC were isolated, and co-cultured with glioma cell line CGH5 in vitro, respectively.The CHG5 cells were set as the control.The effects of GVEC on the migration ability and cytoskeleton of CHG5 cells were analyzed using the Transwell migration assay and immunofluorescence staining, respectively.The expression levels of Rac1 and Cdc42 in CHG5 cells after co-culture were detected using Western blotting. ResultsThe migration ability of CHG5 co-cultured with GVEC was significantly higher than that in CHG5 co-cultured with VEC and CHG5 culture alone (P < 0.05).The results of cytoskeleton immunofluorescence staining showed that the morphology of tubulin in CHG5 cells changed, and the tubulin was thick, circular and radial arrangement after the CHG5 cells were co-cultured with GVEC.The results of Western blotting showed that the expression levels of Rac1 and Cdc42 in CHG5 cells significantly increased after the CHG5 cells were co-cultured with GVEC (P < 0.05), while no obvious change of the expression levels of Rac1 and Cdc42 in CHG5 cells was found after the CHG5 cells were co-cultured with VEC (P>0.05). ConclusionsThe role of tumor cell vascularity is crucial in the migration of glioma cells.The GVEC can promote the migration of tumor cells by changing the cellular tubulin structure, and up-regulating the expression levels of Rac1 and Cdc42 proteins. -
Key words:
- glioma /
- tumor vascular endothelial cell /
- cell migration
-
表 1 不同培养的胶质瘤CHG5细胞迁移能力的比较(x±s)
分组 迁移能力 F P MS组内 CHG5 57.33±2.08 VEC+CHG5 54.00±3.00 1342.28 < 0.01 6.542 GVEC+CHG5 149.33±2.51*# q检验:与CHG5组比较*P < 0.05;与VEC+CHG5组比较#P < 0.05  下载: 导出CSV
下载: 导出CSV
表 2 不同培养的胶质瘤CHG5细胞Rac1、Cdc42蛋白表达的比较(x±s)
分组 Rac1 Cdc42 CHG5 0.907±0.047 0.861±0.108 VEC+CHG5 0.977±0.011 1.113±0.013 GVEC+CHG5 1.156±0.021*# 1.444±0.065*# F 49.90 44.54 P < 0.01 < 0.01 MS组内 0.001 0.006 q检验:与CHG5组比较*P < 0.05;与VEC+CHG5组比较#P < 0.05
下载: 导出CSV
-
[1] WEN PY, KESARI S.Malignant gliomas in adults[J].N Engl J Med, 2008, 359(5):492. doi: 10.1056/NEJMra0708126 [2] HARDEE ME, ZAGZAG D.Mechanisms of glioma-associated neovascularization[J].Am J Pathol, 2012, 181(4):1126. doi: 10.1016/j.ajpath.2012.06.030 [3] SHARMA D, BRUMMEL-ZIEDINS KE, BOUCHARD BA, et al.Platelets in tumor progression:a host factor that offers multiple potential targets in the treatment of cancer[J].J Cell Physiol, 2014, 229(8):1005. doi: 10.1002/jcp.v229.8 [4] DI VITO C, NAVONE SE, MARFIA G, et al.Platelets from glioblastoma patients promote angiogenesis of tumor endothelial cells and exhibit increased VEGF content and release[J].Platelets, 2017, 28(6):585. doi: 10.1080/09537104.2016.1247208 [5] SEGARRA M, KIRCHMAIER BC, ACKER-PALMER A.A vascular perspective on neuronal migration[J].Mech Dev, 2015, 138 Pt 1:17. [6] 韩易, 苏贺先, 马骏, 等.肿瘤血管内皮细胞对胶质瘤细胞侵袭力的影响[J].临床神经外科杂志, 2011, 8(3):139. doi: 10.3969/j.issn.1672-7770.2011.03.009 [7] WATANABE T, NORITAKE J, KAIBUCHI K.Regulation of microtubule in cell migration[J].Trends Cell Biol, 2005, 15(2):76. doi: 10.1016/j.tcb.2004.12.006 [8] LAWSON CD, BURRIDGE K.The on-off relationship of Rho and Rac during integrin-mediated adhesion and cell migration[J].Small GTPases, 2014, 5(1):1. [9] HAGA RB, RIDLEY AJ.Rho GTPases:regulation and roles in cancer cell biology[J].Small GTPases, 2016, 7(4):207. doi: 10.1080/21541248.2016.1232583 -

 点击查看大图
点击查看大图

图(3)表(2)
计量
- 文章访问数: 5132
- HTML全文浏览量: 3318
- PDF下载量: 6
- 被引次数: 0
