




-
肺癌是目前发病率与病死率最高的恶性肿瘤,死亡率高主要是由于绝大部分病人确诊时已属中晚期,5年生存率极低[1]。因此,肺癌的早期诊断和有效的治疗方法是改善预后和提高生存率的关键。近年来,一些研究发现微小RNA(microRNA,miRNA)能调节多种肿瘤的发生、发展及转移等[2]。循环miRNAs可作为肺癌诊断标记物[3],还可在预测肿瘤治疗反应、疗效评价[4]及预后[5]等方面起到重要作用。
miRNA广泛存在于真核生物中,是一种在进化中高度保守的短链非编码RNA[6]。let-7是人体内第一个被发现的miRNA,包含13个成员:let-7-a-1/2/3,let-7-b,let-7-c,let-7-d,let-7-e,let-7-f-1/2,let-7-g,let-7-i,mir-98和mir-202[7]。let-7家族是目前研究最多的miRNA之一,研究[8-10]发现其在多种肿瘤中表达下调,可能作为癌基因或抑癌基因参与肿瘤的发生、发展。关于let-7与肿瘤的研究大多集中在let-7a和let-7b上,关于let-7c的研究较少,而且let-7a1和let-7c共同在肺癌中的研究,至今未见报道。
本研究通过检测肺癌组织、癌旁正常组织和良性肺疾病组织中let-7a1和let-7c基因的表达,分析两者与肺癌发生、发展的相关性,探讨其在肺癌发生中可能的分子机制。
-
本研究获得蚌埠医学院第一附属医院伦理委员会批准,同时标本采集得到病人及家属的知情同意。入组标准:(1)年龄18~75岁;(2)首次确诊的非小细胞肺癌且TNM分期在Ⅲ期以前的病人,或良性肺疾病有外科手术指征者;(3)未行化疗、放疗、靶向治疗及免疫治疗等;(4)肺功能较好能够耐受手术者;(5)无其他手术禁忌证者。排除标准:(1)年龄 < 18岁或>75岁;(2)TNM分期为Ⅳ期的病人,或良性肺疾病无外科手术指征者;(3)接受过化疗、放疗、靶向治疗及免疫治疗等;(4)肺功能较差者;(5)有其他手术禁忌证或拒绝手术治疗者。收取2014年6月至2015年6月在蚌埠医学院第一附属医院胸外科行肺癌根治术的肺癌组织及远离癌组织边缘5 cm的癌旁正常组织44例,其中男25例,女19例,年龄47~72岁,术后病理检查均证实为非小细胞肺癌。其中鳞癌21例,腺癌23例。另外收集良性肺疾病组织9例(肺炎性假瘤3例,结核球2例,肺囊肿1例,复发性气胸1例,支气管扩张1例,胸外伤致左下肺不张1例)作为阴性对照,男8例,女1例,年龄12~57岁。样本一部分放在准备好的RNA保存液中,在4 ℃冰箱过夜后,-80 ℃冰箱保存待用。另一部分行石蜡包埋,病理常规HE染色,观察组织结构。免疫组织化学标记采用SP法,所用一抗TTF1、CK7、NapsinA、p63、p40均购自福州迈新生物技术有限公司,PBS代替一抗作阴性对照。并收集病人吸烟史、肿瘤TNM分期、病理分化程度及有无淋巴结转移等。
-
Trizol试剂(Invitrogen公司);RNA保存液[(天根生化科技(北京)有限公司);DAB显色试剂盒(福州迈新生物技术公司)];EB(北京天根生化科技有限公司);All-in-OneTM miRNA qRT-PCR检测试剂盒(广州复能);let-7a1、let-7c基因引物、U6内参引物(广州复能);SuperScript Ⅲ反转录试剂盒(ABI-invitrogen);Sybr qPCR试剂盒(ABI-invitrogen)。
-
超净工作台(苏州市净化设备有限公司);Finnpipette各型移液器(芬兰ABI-invit rogen);7500型PCR仪(美国ABI公司);AlphaView图像分析软件(美国Alpha公司);Eppendor 5810R型冷冻高速离心机(德国Eppendorf公司);琼脂糖水平电泳槽(美国Bio-Rad公司);MK200-1型干式恒温器(杭州奥盛仪器有限公司);XW-80A型漩涡混合器(上海医科大学仪器厂);VCR76X5型振荡器(海门市其林贝尔仪器制造有限公司)。
-
PCR反应所用的下游引物为试剂盒提供的通用引物,上游引物是广州复能基因提供的All-in-OneTM miRNA qPCR引物;let-7a的上游引物序列号为HmiRQP0002;let-7c的上游引物序列号为HmiRQP0006;U6的上游引物序列号为HmiRQP9001。
-
按说明书进行,用琼脂糖凝胶电泳鉴定总RNA的完整性,28S/18S≥2,说明RNA完整性良好(见图 1)。使用分光光度计测定样品OD值,经紫外分光光度仪分析OD260/OD280,OD260/OD280值均为1.8~2.0。

图 1 总RNA 2%琼脂糖凝胶电泳图
-
染料法荧光定量PCR检测组织中miRNA表达。因成熟let-7很短,只有22 nt,而且其末端不具有mRNA所有的多聚腺苷酸尾巴,所以普通逆转录试剂盒中的随机引物和Olig(dT)引物几乎不能对其进行逆转录。因此,在逆转录前需对其尾端进行加工修饰,本研究选择加尾法修饰进行逆转录,首先利用ploy(A)酶在其尾端进行加尾反应,产物类似mRNA,然后再利用Olig(dT)引物对其进行逆转录,确保荧光定量PCR的特异性。
按All-in-OneTM miRNA qRT-PCR检测试剂盒加尾法逆转录和PCR试剂盒说明书步骤进行,PCR仪器程序参数设置如下:预变性95 ℃、10 min,1个循环;变性95 ℃、10 s,退火60 ℃、24 s,延伸72 ℃、34 s,共40个循环。扩增结束后直接做溶解曲线检测PCR产物的特异性。以cDNA为模板,U6为内对照,检测各标本的let-7a1和let-7c的相对表达水平。
-
50×ROX Reference Dye是荧光定量PCR常用的DNA结合染料。因染料法与双链DNA结合并不是一一对应,一旦有引物二聚体形成或非特异性扩增都将影响结果。所以通过溶解曲线分析判断PCR反应的特异性。最终以内参校正的相对值作为后期统计分析数据。
使用ABI公司的StepOnePlusTM(96孔)实时荧光PCR扩增仪进行实时荧光定量PCR反应,检测各八连管中cDNA的Ct值(C代表荧光信号到达阈值时所需要的PCR反应次数,t代表循环阈值),实验设3个复孔,取平均值,以组织中U6的表达作为内参进行相对定量,采用2-△△Ct值法计算2种miRNA在组织中的相对表达量。公式:△Ct=Ct(let-7)-Ct(U6),△△Ct=△Ct(实验组织)-△Ct(对照组织)。
-
采用t检验、方差分析和q检验。
-
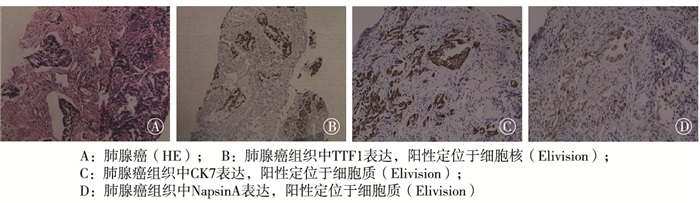
肺腺癌病理显示肿瘤结节状生长,浸润周围肺组织;低倍镜下癌细胞呈不规则腺管状排列,在胶原纤维间浸润性生长。免疫组织化学结果显示癌细胞表达TTF1、CK7、NapsionA(见图 2)。肺鳞癌病理显示不规则结节状生长,与周围肺组织界限不清;低倍镜下癌细胞呈大小不一的巢团状生长,可见小的角化珠及细胞内角化,间质胶原化。免疫组织化学结果显示瘤细胞表达p63、p40(见图 3)。

图 2 肺腺癌HE染色及免疫组织化学标记结果

图 3 肺鳞癌HE染色及免疫组织化学标记结果
-
荧光定量PCR扩增曲线呈较为光滑的S型,均在30个循环以前出现扩增信号,结果较为可靠;由于产物序列不同,溶解曲线荧光信号主峰基本集中在80~90 ℃和78~88 ℃,其前有小的杂峰,考虑为复空中底物浓度过低所至引物二聚体的形成(见图 4~7)。

图 4 let-7a1扩增曲线Line图

图 5 let-7c扩增曲线Line图

图 6 内参U6扩增曲线Line图

图 7 三者溶解曲线
-
肺癌组织中let-7a1和let-7c的相对表达量均低于癌旁正常组织和良性肺疾病组织(P < 0.05);癌旁正常组织中let-7a1和let-7c的相对表达量均低于良性肺疾病组织(P < 0.05)(见表 1)。
分组 n let-7a1 let-7c 肺癌组织 44 0.93± 0.46*# 1.16± 0.76*# 癌旁正常组织 44 1.18±0.54* 1.59±0.88* 良性肺疾病组织 9 1.38±0.54 1.81±0.63 F — 4.39 4.30 P — < 0.05 < 0.05 MS组内 — 0.026 0.652 q检验:与良性肺疾病组织比较*P < 0.05;与癌旁组织比较#P < 0.05 表 1 肺癌组织、癌旁正常组织和良性肺疾病组织中let-7a1、let-7c基因的表达比较(x±s)
-
let-7a和let-7c基因在肺鳞癌和腺癌中的相对表达量差异均无统计学意义(P>0.05)(见表 2);在肺鳞癌和腺癌旁正常组织中的相对表达量差异也均无统计学意义(P>0.05)(见表 3)。
分组 n let-7a1 let-7c 鳞癌 21 1.01±0.41 1.02±0.83 腺癌 23 0.86±0.49 1.07±0.90 t — 1.12 0.86 P — >0.05 >0.05 表 2 let-7a1和let-7c基因在不同病理类型肺癌组织中的表达比较(x±s)
分组 n let-7a1 let-7c 鳞癌旁正常组织 21 1.33±0.62 1.61±0.74 腺癌旁正常组织 23 1.05±0.41 1.57±1.00 t — 1.78 0.16 P — >0.05 >0.05 表 3 let-7a和let-7c基因在不同病理类型肺癌旁正常组织中的表达比较(x±s)
-
let-7a1表达与分化程度、吸烟和TNM分期有关(P < 0.05~P < 0.01),与淋巴结有无转移无关(P>0.05)。let-7c表达与淋巴结转移、分化程度有关(P < 0.01),与吸烟、TNM分期无关(P>0.05)(见表 4)。
临床特征 n let-7a1 let-7c 淋巴结转移 有 30 0.90±0.43 0.80±0.31 无 14 1.00±0.52 1.95±0.86 t — 0.66 6.54 P — >0.05 < 0.01 分化程度 低 4 0.35±0.11 1.04±0.30 中 27 0.79±0.28* 0.92±0.51 高 13 1.42±0.39*# 1.71±1.02# F — 26.58 5.85 P — < 0.01 < 0.01 吸烟 有 23 0.63±0.21 1.04±0.52 无 21 1.27±0.41 1.29±0.96 t — 6.64 1.08 P — < 0.01 >0.05 TNM分期 ≤Ⅲa期 39 0.99±0.45 1.20±0.80 Ⅲb期 5 0.50±0.28 0.87±0.38 t — 2.41 0.91 P — < 0.05 >0.05 q检验:与低分化组比较*P < 0.05;与中分化组比较#P < 0.05 表 4 let-7a-1和let-7c基因与各临床相关指标之间的关系(x±s)
-
肺癌是发病率及死亡率最高的癌症,2018年全球肺癌新发病例达到209万余例,肺癌相关死亡病例高达176万余例,同年我国统计数据显示约有77.4万的新增肺癌病例,其中69万人死于肺癌[11]。非小细胞肺癌占肺癌的80%~90%,而小细胞癌在过去20多年里在许多国家的发病率一直在下降[12]。对于非小细胞肺癌,近年来尽管采用了手术、化疗、放疗、靶向治疗和免疫治疗等,其5年生存率仅15%[13]。靶向和免疫治疗一直是肺癌领域研究的热点,但也存在较多问题,靶向治疗主要针对有EGFR、ALK、c-MET、ROS1等突变的病人,但仍有大约有一半的非小细胞肺癌病人无明确靶点,同时靶向治疗耐药不可避免;免疫治疗开创肿瘤治疗新纪元,一旦获益可能长期生存,但获益率仅20%左右[14],且有一定不良反应。因此,进一步阐明肺癌发展演变过程中分子机制,提高靶向和免疫治疗的疗效将对治疗肺癌具有重大意义。
本研究通过荧光定量PCR检测肺癌组织中let-7a1、let-7c的表达情况并结合临床资料,分析其可能的分子机制。结果显示,肺癌组织中let-7a1和let-7c基因表达低于癌旁组织和正常肺组织,差异有统计学意义;但在腺癌、鳞癌组织及其癌旁组织中表达,差异均无统计学意义;这与尚学芹等[15]研究结果一致,也与其在其他肿瘤组织中的表达结果一致[16-17]。但FASSINA等[18]研究认为,在腺癌组织中let-7a、let-7b、let-7c、let-7f、let-7g、let-7i和miR-98的表达量明显高于鳞癌组织,考虑本研究所使用的均为手术标本,均经严格的标本清洗及储存处理,而且本研究样本量44例高于FASSINA等研究的所采用的针吸标本组织31例,故认为本研究结果更可信。
本研究结果显示,Let-7c在肺癌组织中的表达与淋巴结转移、分化程度有关,与ZHAO等[19]的报道一致,也与耿淼等[20]在乳腺癌和ZHU等[21]在肝癌细胞中体外及体内实验结果一致,但具体作用机制有待进一步研究。ZHAO等[19]认为肺癌的转移与let-7c下游靶基因整合素β3和丝裂原蛋白激酶激酶激酶激酶3有关,这给肺癌的淋巴结转移途径研究提供新的证据。对于本研究发现的let-7c与吸烟、TNM分期无关,尚未见文献报道,其发生机制我们将进一步探讨。与之不同的是let-7a1则与吸烟、TNM分期明显相关,而与淋巴结转移无关。另外let-7a1在肺癌中的表达随分化程度越差其表达明显减低。同时本研究结果还显示,let-7a1表达在一定程度上与吸烟相关,有吸烟史者表达减低,这与IZZZOTTI等[22]研究的烟草暴露环境中小鼠的肺组织上皮细胞let-7表达下降的结果基本一致。
目前关于let-7在肺癌中的治疗价值主要集中在以下几方面:(1)含铂两药方案一直是肺癌的标准化疗方案,但有效率较低,客观缓解率为29.2%~36.4%[23]。最近有研究[24]认为肺癌细胞对铂类药物的耐药可能也与let-7和miR-29对Cockayne syndrome protein B的调节有关。(2)针对非小细胞肺癌中上调let-7基因的研究认为高迁移率族蛋白A2 mRNA是let-7b的功能性小分子RNA,在HCC827细胞的的体内外研究结果都表明能使其特异性沉默,因此认为let-7可能是潜在的肿瘤治疗靶点[25]。另有研究[26]发现miR-203通过抑制LIN28B和促进let-7的生物合成,在抑制肺癌细胞增殖和促进凋亡方面发挥了重要作用。(3)靶向治疗一直是非小细胞肺癌治疗的热点,特别是EGFR-TKIs的临床应用,给病人带来了无进展生存期的获益,改善了生活质量,但耐药不可避免,其分子机制尚不清楚。有研究[27]报道与吉非替尼敏感PC9细胞相比,吉非替尼耐药PC9/GR细胞let-7家族表达下调,miR-17家族表达上调。let-7和miR-17参与了非小细胞肺癌EGFR-TKI耐药的调控,可作为预测非小细胞肺癌EGFR-TKI耐药的生物标志物。(4)免疫检查点抑制剂是肿瘤治疗的一个最有前途的靶点,但客观应答率小于40%,并且对PD-1/PD-L1表达调控的机制知之甚少。CHEN等[28]研究发现let-7能够抑制PD-L1表达,而LIN28是一种RNA结合蛋白,在大多数癌细胞中上调,抑制let-7的生物合成,从而促进PD-L1的表达。因此,抑制LIN28可能是预防癌细胞免疫逃逸的一种策略。因此let-7已成为肿瘤治疗和免疫应答的关键分子,本身也具有潜在的免疫治疗靶点作用[29]。
LI等[30]通过Meta分析探讨microRNA let-7在肺癌中的预后作用也发现let-7的低表达提示整体生存率低,危险比为1.55(95%CI:1.16~2.09,P < 0.05)。let-7的低表达与肺癌病人预后不良密切相关。
综上,let-7a1和let-7c基因在肺癌组织中低表达,并且随临床分期进展、淋巴结转移呈明显下降趋势,提示两者与肺癌的发生、发展及转移密切相关,let-7a1和let-7c可能成为肺癌早期诊断生物标记因子,并可能成为肺癌的靶向治疗新的靶点,为高危人群的早期干预提供实验依据。
let-7a1和let-7c在非小细胞肺癌中的表达及临床意义
Expression and clinical significance of let-7a1 and let-7c in non-small cell lung cancer
-
摘要:
目的探讨人类微小RNA let-7a1和let-7c在肺癌组织中的表达及与临床病理特征之间的关系。 方法收集53例肺组织手术标本及临床资料,其中包括肺癌组织44例(收集其癌组织和癌旁正常组织)和良性肺疾病组织9例。并收集病人吸烟史、肿瘤TNM分期、病理分化程度及有无淋巴结转移等。采用Trizol法提取总RNA,采用荧光定量PCR检测let-7a1和let-7c的表达,分析其在肺癌组织、癌旁正常组织及良性肺疾病组织中的表达差异及与临床特征的关系。 结果肺癌组织中let-7a1和let-7c的相对表达量均低于癌旁正常组织和良性肺疾病组织(P < 0.05);癌旁正常组织中let-7a1和let-7c的相对表达量均低于良性肺疾病组织(P < 0.05)。let-7a1表达与分化程度、吸烟和TNM分期有关(P < 0.05~P < 0.01),与淋巴结有无转移无关(P>0.05)。let-7c表达与淋巴结转移、分化程度有关(P < 0.01),与吸烟、TNM分期无关(P>0.05)。 结论let-7a1和let-7c基因在肺癌组织中相对表达量降低,并且随临床分期进展、淋巴结转移呈明显下降趋势,提示两者与肺癌的发生、发展及转移密切相关,let-7a1和let-7c可能成为肺癌早期诊断生物标志因子,并可能成为肺癌的靶向治疗新的靶点,为高危人群的早期干预提供实验依据。 Abstract:ObjectiveTo investigate the expression of human microRNA let-7a1 and let-7c in lung cancer tissues and its relationship with clinicopathological features. MethodsThe surgical specimens and clinical data of 53 cases of lung tissue were collected, including 44 cases of lung cancer used to collect cancer tissue and paracancerous normal tissue, and 9 cases of benign lung disease tissue.The smoking history, TNM stage, pathological differentiation and lymph node metastasis were collected.The total RNA was extracted by Trizol method, and the expression of let-7a1 and let-7c was detected by fluorescence quantitative PCR.The difference of let-7a1 and let-7c expression in lung cancer tissue, paracancerous normal tissue and benign lung disease tissue, and its relationship with clinical features were analyzed. ResultsThe relative expression of let-7a1 and let-7c in lung cancer tissue was lower than that in paracancerous normal tissue and benign lung disease tissue(P < 0.05), and the relative expression of let-7a1 and let-7c in paracancerous normal tissue was lower than that in benign lung disease tissue(P < 0.05).The expression of let-7a1 was related to differentiating degree, smoking and TNM stage (P < 0.05 to P < 0.01), but was not related to smoking and TNM stage (P>0.05).The expression of let-7c was related to lymph node metastasis and differentiating degree (P < 0.01), but was not related to smoking and TNM stage (P>0.05). ConclusionsThe relative expression of let-7a1 and let-7c genes in lung cancer tissue decreases, and exhibits decreasing trend with the progress of clinical stage and lymph node metastasis, which suggests that let-7a1 and let-7c are closely related to the occurrence, development and metastasis of lung cancer.Let-7a1 and let-7c may become biomarkers for early diagnosis of lung cancer, and become new targets for targeted therapy of lung cancer, to provide experimental basis for early intervention of high-risk groups. -
Key words:
- lung neoplasms /
- microRNA /
- let-7a1 /
- let-7c
-
表 1 肺癌组织、癌旁正常组织和良性肺疾病组织中let-7a1、let-7c基因的表达比较(x±s)
分组 n let-7a1 let-7c 肺癌组织 44 0.93± 0.46*# 1.16± 0.76*# 癌旁正常组织 44 1.18±0.54* 1.59±0.88* 良性肺疾病组织 9 1.38±0.54 1.81±0.63 F — 4.39 4.30 P — < 0.05 < 0.05 MS组内 — 0.026 0.652 q检验:与良性肺疾病组织比较*P < 0.05;与癌旁组织比较#P < 0.05  下载: 导出CSV
下载: 导出CSV
表 2 let-7a1和let-7c基因在不同病理类型肺癌组织中的表达比较(x±s)
分组 n let-7a1 let-7c 鳞癌 21 1.01±0.41 1.02±0.83 腺癌 23 0.86±0.49 1.07±0.90 t — 1.12 0.86 P — >0.05 >0.05
下载: 导出CSV
表 3 let-7a和let-7c基因在不同病理类型肺癌旁正常组织中的表达比较(x±s)
分组 n let-7a1 let-7c 鳞癌旁正常组织 21 1.33±0.62 1.61±0.74 腺癌旁正常组织 23 1.05±0.41 1.57±1.00 t — 1.78 0.16 P — >0.05 >0.05
下载: 导出CSV
表 4 let-7a-1和let-7c基因与各临床相关指标之间的关系(x±s)
临床特征 n let-7a1 let-7c 淋巴结转移 有 30 0.90±0.43 0.80±0.31 无 14 1.00±0.52 1.95±0.86 t — 0.66 6.54 P — >0.05 < 0.01 分化程度 低 4 0.35±0.11 1.04±0.30 中 27 0.79±0.28* 0.92±0.51 高 13 1.42±0.39*# 1.71±1.02# F — 26.58 5.85 P — < 0.01 < 0.01 吸烟 有 23 0.63±0.21 1.04±0.52 无 21 1.27±0.41 1.29±0.96 t — 6.64 1.08 P — < 0.01 >0.05 TNM分期 ≤Ⅲa期 39 0.99±0.45 1.20±0.80 Ⅲb期 5 0.50±0.28 0.87±0.38 t — 2.41 0.91 P — < 0.05 >0.05 q检验:与低分化组比较*P < 0.05;与中分化组比较#P < 0.05
下载: 导出CSV
-
[1] TORRE LA, SIEGEL RL, JEMAL A. Lung cancer statistics[J]. Adv Exp Med Biol, 2016, 893: 1. [2] REDDY KB. MicroRNA(miRNA) in cancer[J]. Cancer Cell Int, 2015, 15(1): 38. doi: 10.1186/s12935-015-0185-1 [3] WANG C, DING M, XIA M, et al. A five-miRNA panel identified from a multicentric case-control study serves as a novel diagnostic tool for ethnically diverse non-small-cell lung cancer patients[J]. Bio Medicine, 2015, 2(10): 1377. doi: 10.1016/j.ebiom.2015.07.034 [4] FRANCHINA T, AMODEO V, BRONTE G, et al. Circulating miR-22, miR-24 and miR-34a as novel predictive biomarkers to pemetrexed-based chemotherapy in advanced non-small cell lung cancer[J]. J Cell Physiol, 2014, 229(1): 97. [5] MATIKAS A, SYRIGOS KN, AGELAKI S. Circulating biomarkers in non-small-cell lung cancer: current status and future challenges[J]. Clin Lung Cancer, 2016, 17(6): 507. doi: 10.1016/j.cllc.2016.05.021 [6] STEFANI G, SLACK FJ. Small non-coding RNAs in animal development[J]. Nat Rev Mol Cell Bio, 2008, 9(3): 219. doi: 10.1038/nrm2347 [7] BOYERINAS B, PARK SM, HAU A, et al. The role of let-7 in cell differentiation and cancer[J]. Endocrine-Related Cancer, 2010, 17(1): F19. doi: 10.1677/ERC-09-0184 [8] YE Y, LI X, YANG Q, et al. The role of microRNA in human lung squamous cell carcinoma[J]. Cancer Genet Cytogenet, 2010, 200(2): 127. doi: 10.1016/j.cancergencyto.2010.03.014 [9] ZHONG Z, DONG Z, YANG L, et al. Inhibition of proliferation of human lung cancer cells by green tea catechins is mediated by upregulation of let-7[J]. Exp Ther Med, 2012, 4(2): 267. doi: 10.3892/etm.2012.580 [10] ZHAN M, QU Q, WANG G, et al. Let-7c inhibits NSCLC cell proliferation by targeting HOXA1[J]. APJCP, 2013, 14(1): 387. doi: 10.7314/APJCP.2013.14.1.387 [11] 中国临床肿瘤学会血管靶向治疗专家委员会, 非小细胞肺癌专家委员会, 非小细胞肺癌抗血管生成药物治疗专家组. 晚期非小细胞肺癌抗血管生成药物治疗中国专家共识(2019版)[J]. 中国肺癌杂志, 2019, 22(7): 401. [12] BRAY F, FERLAY J, SOERJOMATARAM I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2018, 68(6): 394. doi: 10.3322/caac.21492 [13] 何圆, 尤长宣. 非小细胞肺癌免疫治疗进展[J]. 中国肺癌杂志, 2014, 17(3): 277. [14] 张娣, 黄架旗, 张初峰, 等. PD-1/PD-L1免疫检查点抑制剂在肺癌中的研究进展[J]. 中国肺癌杂志, 2019, 22(6): 366. [15] 尚学琴, 阎婷婷, 杨眉, 等. 肺癌组织中Let-7的表达及意义[J]. 广东医学, 2018, 39(7): 983. [16] 董峰, 马君俊, 孙晶, 等. miR-124a、let-7表达与胃癌临床病理特征和化疗疗效的相关性[J]. 现代生物医学进展, 2018, 18(24): 4643. [17] 喻箴, 肖明耀, 崔明, 等. Let-7在正常乳腺组织和乳腺癌组织中的表达[J]. 昆明医科大学学报, 2016, 37(7): 19. [18] FASSINA A, CAPPELLESSO R, FASSAN M. Classification of non-small cell lung carcinoma in transthoracic needle specimens using microRNA expression profiling[J]. Chest, 2011, 140(5): 1305. doi: 10.1378/chest.11-0708 [19] ZHAO B, HAN H, CHEN J, et al. MicroRNA let-7c inhibits migration and invasion of human non-small cell lung cancer by targeting ITGB3 and MAP4K3[J]. Cancer Letters, 2014, 342(1): 43. doi: 10.1016/j.canlet.2013.08.030 [20] 耿淼, 潘静坤, 罗芸, 等. miRNA Let-7与乳腺癌预后的相关性分析[J]. 局解手术学杂志, 2019, 28(1): 1. [21] ZHU X, WU L, YAO J, et al. MicroRNA let-7c inhibits cell proliferation and induces cell cycle arrest by targeting CDC25A in human hepatocellular carcinoma[J]. PLoS One, 2015, 10(4): e0124266. doi: 10.1371/journal.pone.0124266 [22] IZZOTTI A, CALIN GA, ARRIGO P, et al. Downregulation of microRNA expression in the lungs of rats exposed to cigarette smoke[J]. Faseb J, 2009, 23(3): 806. doi: 10.1096/fj.08-121384 [23] 李玉梅, 刘静, 周争光, 等. 培美曲塞与吉西他滨联合顺铂一线治疗晚期非小细胞肺癌的对比研究[J]. 蚌埠医学院学报, 2018, 43(6): 719. [24] YANG Z, LIU C, WU H, et al. CSB affected on the sensitivity of lung cancer cells to platinum-based drugs through the global decrease of let-7 and miR-29[J]. BMC Cancer, 2019, 19(1): 948. doi: 10.1186/s12885-019-6194-z [25] MEIRAV S, ANNABELLE B, MAUD-EMMANUELLE G, et al. Hydrophobically modified let-7b miRNA enhances biodistribution to NSCLC and downregulates HMGA2 in vivo[J]. Mol Ther Nucleic Acids, 2020, 19: 267. doi: 10.1016/j.omtn.2019.11.008 [26] ZHOU Y, LIANG H, LIAO Z, et al. MIR-203 enhances let-7 biogenesis by targeting LIN28B to suppress tumor growth in lung cancer[J]. Sci Rep, 2017, 7: 42680. doi: 10.1038/srep42680 [27] YIN J, HU W, PAN L, et al. Let-7 and miR-17 promote self-renewal and drive gefitinib resistance in non-small cell lung cancer(Article)[J]. Oncol Rep, 2019, 42(2): 495. doi: 10.3892/or.2019.7197 [28] CHEN Y, XIE C, ZHENG X, et al. LIN28/let-7/PD-L1 pathway as a target for cancer immunotherapy[J]. Cancer Immunol Res, 2019, 7(3): 487. doi: 10.1158/2326-6066.CIR-18-0331 [29] GILLES ME, SLACK FJ. Let-7 microRNA as a potential therapeutic target with implications for immunotherapy[J]. Expert Opin Ther Targets, 2018, 22(11): 929. doi: 10.1080/14728222.2018.1535594 [30] LI XX, DI X, CONG S, et al. The role of let-7 and HMGA2 in the occurrence and development of lung cancer: a systematic review and meta-analysis[J]. Eur Rev Med Pharmacol Sci, 2018, 22(23): 8353. doi: 10.26355/eurrev_201812_16533 -

 点击查看大图
点击查看大图

图(7)表(4)
计量
- 文章访问数: 4832
- HTML全文浏览量: 2631
- PDF下载量: 58
- 被引次数: 0
