


-
丙型肝炎病毒(hepatitis C virus, HCV)可导致慢性肝脏损伤、肝纤维化、肝硬化甚至癌变可能。HCV非结构蛋白4B(non-structural protein 4B, NS4B)是HCV编码的非结构蛋白之一,其功能虽未完全明了,但已有实验证据表明NS4B是参与HCV复制[1]、组装[2]和致病的重要成分。研究[3]发现NS4B诱导细胞的内质网应激和错误折叠蛋白反应,使大量位于内质网上HCV编码的蛋白有效折叠,从而在有利于组装HCV有效复制复合物的同时又不引起细胞凋亡[4]。本课题组前期研究[5]发现,HCV NS4B可通过泛素化途径参与宿主细胞癌变过程;且我们的最近研究[6]证实,NS4B可通过神经纤维瘤病Ⅱ型(neurofibromatosis type 2, NF2)(所表达蛋白为Merlin)调节细胞周期,但具体机制不明。本研究针对我们的前期研究成果,通过检测NS4B对NF2下游Hippo信号通路的重要因子Yes相关蛋白(Yes associated protein, YAP)转录、表达、磷酸化水平等的影响,探讨NS4B影响细胞周期的可能机制,以期为HCV的致病机制提供参考。
-
质粒转染试剂lipofectamineTM2000(Life technology公司);YAP、P-YAP、NS4B、Lamin、Tubulin等抗体(Abcam公司);总RNA提取试剂、逆转录试剂盒、质粒提取试剂盒、普通PCR反应试剂盒以及Real-time PCR试剂盒等(Omega公司)。
-
NS4B PCR扩增引物序列见表 1。Real-time PCR引物序列见表 2。
基因 正向引物(5′-3′) 反向引物(5′-3′) NS4B GGA ATT CTA TGG CCG CAC ACC GCT CTA GAA GTG GAC AAG CCT GCT AA TCC CTT ACA TC GCT AA 表 1 NS4B PCR扩增引物
基因 正向引物(5′-3′) 反向引物(5′-3′) hGAPDH CAA GAA GGT GGT GAA GCA GG AAA GGT GGA GGA GTG GGT GT YAP GAG ATC CCT GAT GAT GTA CCA C TGT TGT TGT CTG ATC GTT GTG AT 表 2 Real-time PCR引物
-
pCDNA3.1、pCDNA3.1-NS4B及pEGFPC1质粒(清华大学深圳研究生院黄来强教授实验室馈赠)。pCMV2-NF2质粒、HEPG2和293T细胞系均由本实验室保存。
-
NS4B完整序列经扩增酶切后与荧光质粒pEGFPC1连接,重组质粒经酶切测序验证后,转染进入HEPG2和293T细胞内,通过Western blotting验证目的蛋白表达效果;通过免疫荧光验证荧光蛋白表达效果。
-
4组HEPG2细胞,其中2组各转染2 μg pCDNA3.1-NS4B,另外2组各转染2 μg pCDNA3.1,继续培养至24 h和48 h时点各取一组收样,检测细胞内YAP转录、表达及磷酸化水平;不同浓度NF2表达质粒和Si-NF2 RNA序列[6]分别转入8份HEPG2细胞继续培养24 h后收样,检测细胞内YAP蛋白磷酸化水平。
-
细胞爬片,将荧光表达质粒pEGFPC1-NS4B及对照质粒pEGFPC1各2 μg分别转入293T细胞内,24 h后收获细胞进行免疫荧光检测。
-
待细胞密度为80%左右时,倒去培养液,预冷PBS洗2次,用胰蛋白酶消化细胞,进行细胞计数,以每孔(0.5~1.0)×106进行铺板(6孔板),培养过夜,细胞增殖到约80%密度时进行转染。严格按照lipofectamineTM2000说明书内容进行质粒转染。质粒转染完成后于继续培养的24 h和48 h时间点收获细胞,提取总RNA及蛋白,分别进行Real-time PCR和Western blotting检测。
-
pEGFPC1-NS4B及pEGFPC1质粒各2 μg分别转入293T细胞24 h后,弃去培养液,用PBS轻柔洗涤细胞2~3次,4%多聚甲醛固定细胞10~15 min;PBS轻柔洗涤细胞2~3次,5分钟/次,10% Triton-100打孔5 min,再用PBS轻柔洗涤细胞2~3次,5分钟/次,5%胎牛血清封闭2 h。YAP蛋白一抗孵育过夜。PBS轻柔洗涤细胞2~3次,5分钟/次,二抗遮光孵育2 h,PBS洗涤细胞2~3次,5分钟/次,封片剂封片,激光共聚焦显微镜下观察,拍照。
-
4组HEPG2细胞,其中2组各转染2 μg pCDNA3.1-NS4B,另外2组各转染2 μg pCDNA3.1,继续培养至24 h和48 h时点各取一组收样,置于1.5 mL EP管内,离心,去上清液。使用博士德胞质胞核蛋白提取试剂盒,对收集细胞进行细胞核质分离并进行Western blotting检测。
-
RNA反转录、cDNA合成、Real-time PCR反应、质粒提取、Western blotting等均严格按照试剂说明书或相关参考书进行。
-
采用t检验。
-
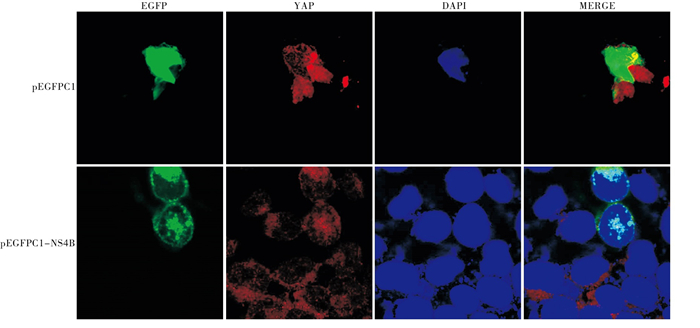
pEGFPC1-NS4B重组质粒经酶切电泳后可观察到目的基因大小一致条带(见图 1);测序结果经在NCBI基因库比对后显示为完整HCV 1b NS4B基因序列;HEPG2细胞内转入pEGFPC1-NS4B质粒24 h收集细胞提取蛋白,使用NS4B特异性抗体进行Western blotting检测可检测到相应蛋白;pEGFPC1-NS4B质粒转染293T细胞24 h后,荧光显微镜下可观察到绿色荧光信号(见图 2)。

图 1 pEGFPC1-NS4B的构建

图 2 pEGFPC1-NS4B的表达检测
-
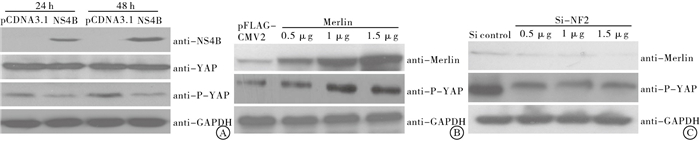
NS4B转入HEPG2细胞内24、48 h,YAP转录水平均与对照组差异均无统计学意义(P>0.05)(见表 1);但YAP磷酸化水平明显改变,且在HEPG2细胞内高表达NF2后,YAP磷酸化水平升高;反之,HEPG2细胞内低表达NF2后,YAP磷酸化水平降低(见图 3)。
分组 n 24 h 48 h pCDNA3.1 3 1.00±0.03 1.00±0.01 pCDNA3.1-NS4B 3 0.96±0.05 1.08±0.05 t — 1.15 2.41 P — >0.05 >0.05 表 1 NS4B对YAP转录影响(x±s)

图 3 NS4B及NF2对Y AP转录和表达影响
-
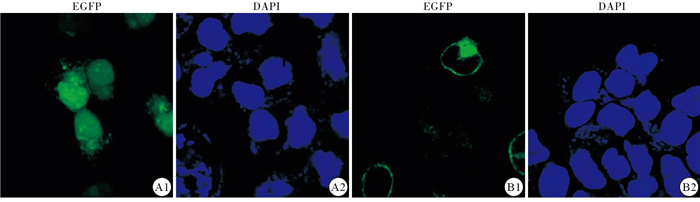
与pEGFPC1空质粒对照组比较,pEGFPC1-NS4B转入293T 48 h后明显使YAP细胞内分布发生改变。与对照组YAP在细胞质和细胞核内分布比较,pEGFPC1-NS4B组YAP在细胞核内的分布明显增多(见图 4)。另外,在HEPG2高表达NS4B后进行核质分离试验,发现高表达NS4B后细胞质内的YAP总量减少,细胞核内的YAP总量升高,并且P-YAP在细胞质内的含量减少,与激光共聚焦试验结果一致(见图 5)。

图 4 激光共聚焦试验检测293T细胞内高表达NS4B后YAP蛋白定位

图 5 Western-blotting检测HEPG2细胞内高表达NS4B后细胞核及细胞质内YAP和P-YAP含量
-
Hippo信号通路最初在果蝇遗传学研究中发现,目前认为Hippo通路主要和细胞增殖、凋亡、器官组织生长等方面发挥关键作用[7-8]。YAP蛋白是Hippo途径的关键蛋白,其主要生物学功能和细胞的增殖及凋亡相关[9-10]。YAP主要在细胞核内发挥生物学作用,而是否能够进入细胞核内则主要由YAP的磷酸化程度决定。磷酸化后的YAP滞留在细胞质中,无法进入细胞核,从而失活。而其磷酸化水平主要受Hippo途径上游因子例如NF2蛋白的调控。
本课题组前期研究[6]发现,HCV NS4B能够通过NF2调节细胞周期。Merlin蛋白是PI3K/Akt[11]、Hippo[12-13]等多个信号通路的调节因子,与细胞周期调控关系密切,主要表现在抑制细胞异常增殖,增加G0/G1期细胞比例[14-15],此外还与细胞膜稳定性、细胞信号传递及接触抑制有关。HCV NS4B是否通过NF2激活了其下游信号通路使宿主细胞发生生物学改变,这一点仍不明了。基于此,本研究以Hippo信号通路的YAP蛋白为靶因子,重点探讨NS4B对YAP转录、表达、磷酸化及细胞内分布影响,以逐步揭示NS4B调节细胞周期的具体机制。
本研究构建了NS4B的荧光质粒pEGFPC1-NS4B,以观察NS4B和YAP蛋白细胞内定位。通过菌落PCR、酶切鉴定及测序结果证实插入pEGFPC1质粒的基因片段为完整的NS4B表达序列。将pEGFPC1和pEGFPC1-NS4B分别转入2组293T细胞内,通过共聚焦显微镜观察可以发现,pEGFPC1质粒组的荧光蛋白散在分布整个细胞内,而pEGFPC1-NS4B的荧光蛋白则主要呈团块状分布;另外,Western blotting结果显示,pEGFPC1-NS4B转入HEPG2细胞24 h后可以检测到与NS4B特异性抗体结合的蛋白,提示pEGFPC1-NS4B质粒构建成功。
本研究转入NS4B质粒后24、48 h时YAP转录及表达水平与空质粒pCDNA3.1组相比均无明显差异,说明在HEPG2细胞内NS4B并不影响YAP的转录和表达;但是YAP的磷酸化水平却明显降低;同时,通过高表达和低表达NF2后检测YAP磷酸化水平,结果显示,高表达NF2可上调YAP的磷酸化水平,相反,低表达NF2后可上调YAP的磷酸化水平。结合实验组前期研究[6]成果,提示NS4B可通过NF2调节Hippo通路,从而发挥生物学功能。以往研究[9-10]证实,YAP磷酸化会使YAP蛋白失活,使YAP滞留在细胞质中,从而导致细胞的增殖减缓或引起凋亡发生,而YAP去磷酸化则激活YAP蛋白活性,从而发挥去促进细胞增殖和抑制凋亡的功能。激光共聚焦和核质分离实验发现NS4B转入细胞后能够促进YAP蛋白在细胞核内的分布,与以往研究相一致。提示NS4B可能通过影响YAP磷酸化从而调节细胞增殖和凋亡,但这一点仍需要进行细胞增殖及凋亡的相关实验加以验证。鉴于肿瘤细胞增殖不受限和凋亡减弱的特性,可推测YAP或Hippo信号通路可能参与HCV导致肝脏癌变的机制。
综上,本研究成功构建了HCV NS4B的荧光载体质粒,并发现NS4B对Hippo通路关键因子YAP转录、表达、磷酸化及细胞内分布影响,初步验证了NS4B对NF2下游通路的影响,为进一步揭示NS4B调节细胞周期及增殖的机制提供思路。
丙型肝炎病毒NS4B对YAP转录、表达、磷酸化及细胞内分布的影响
Effect of the hepatitis C virus non-structural protein 4B on the levels of YAP transcription, expression, phosphorylation and intracellular distribution
-
摘要:
目的 构建丙型肝炎病毒(HCV)非结构蛋白4B(NS4B)细胞内荧光表达质粒,检测NS4B对Yes相关蛋白(YAP)转录、表达、磷酸化及细胞内分布影响。 方法根据现有含有HCV 1b型病毒NS4B完整基因的质粒设计NS4B的特异性扩增引物,扩增NS4B完整序列并构建NS4B荧光质粒,质粒转染进入HEPG2和293T细胞,通过Real-time PCR检测YAP基因转录水平;通过Western blotting检测YAP蛋白表达及磷酸化水平;通过激光共聚焦和核质分离实验检测YAP细胞内分布。 结果成功扩增出NS4B完整序列并构建其荧光质粒;Real-time PCR和Western blotting结果显示,NS4B并不影响YAP的转录和表达水平,但是能够调节其磷酸化水平;激光共聚焦和核质分离试验结果提示NS4B能够调节YAP在细胞质和细胞核内的比例。 结论HCV NS4B可能通过对YAP的影响发挥生物学功能。 Abstract:ObjectiveTo construct the intracellular fluorescent expression plasmid of non-structural protein 4B(NS4B) of hepatitis C virus(HCV), and detect the effects of NS4B on the levels of transcription, expression, phosphorylation and intracellular distribution of Yes-associated protein(YAP). MethodsThe specific amplification primers for NS4B were designed according to the existing plasmid containing HCV 1b virus NS4B complete gene.The complete sequence of NS4B was amplified to construct the NS4B fluorescent plasmid, and the plasmid was transfected into HEPG2 and 293T cells.The transcription level of YAP gene, and expression and phosphorylation levels of YAP protein were detected using Real-time PCR and Western blotting, respectively.The intracellular distribution of YAP was detected by confocal laser and nucleo-cytoplasmic separation assay. ResultsThe complete NS4B sequence was successfully amplified, and its fluorescent plasmid was constructed.The results of Real-time PCR and Western blotting showed that the NS4B did not affect the transcription and expression levels of YAP, but could regulate its phosphorylation level.The results of laser confocal and nucleo-cytoplasmic separation results suggested that the NS4B could regulate the ratio of YAP in cytoplasm and nucleus. ConclusionsThe HCV NS4B may play a biological role through its effects on YAP. -
表 1 NS4B PCR扩增引物
基因 正向引物(5′-3′) 反向引物(5′-3′) NS4B GGA ATT CTA TGG CCG CAC ACC GCT CTA GAA GTG GAC AAG CCT GCT AA TCC CTT ACA TC GCT AA  下载: 导出CSV
下载: 导出CSV
表 2 Real-time PCR引物
基因 正向引物(5′-3′) 反向引物(5′-3′) hGAPDH CAA GAA GGT GGT GAA GCA GG AAA GGT GGA GGA GTG GGT GT YAP GAG ATC CCT GAT GAT GTA CCA C TGT TGT TGT CTG ATC GTT GTG AT
下载: 导出CSV
表 1 NS4B对YAP转录影响(x±s)
分组 n 24 h 48 h pCDNA3.1 3 1.00±0.03 1.00±0.01 pCDNA3.1-NS4B 3 0.96±0.05 1.08±0.05 t — 1.15 2.41 P — >0.05 >0.05
下载: 导出CSV
-
[1] ZHANG Y, ZOU J, ZHAO X, et al. Hepatitis C virus NS5A inhibitor daclatasvir allosterically impairs NS4B-involved protein-protein interactions within the viral replicase and disrupts the replicase quaternary structure in a replicase assembly surrogate system[J]. J Gen Virol, 2019, 100(1): 69. doi: 10.1099/jgv.0.001180 [2] PAUL D, MADAN V, RAMIREZ O, et al. Glycine zipper motifs in hepatitis C virus nonstructural protein 4B are required for the establishment of viral replication organelles[J]. J Virol, 2018, 92(4): e01890. [3] KONG L, LI S, HUANG M, et al. The roles of endoplasmic reticulum overload response induced by HCV and NS4B protein in human hepatocyte viability and virus replication[J]. PLoS One, 2015, 10(4): e123190. [4] LI S, YE L, YU X, et al. Hepatitis C virus NS4B induces unfolded protein response and endoplasmic reticulum overload response-dependent NF-κB activation[J]. Virology, 2009, 391(2): 257. doi: 10.1016/j.virol.2009.06.039 [5] HU B, LI S, ZHANG Z, et al. HCV NS4B targets Scribble for proteasome-mediated degradation to facilitate cell transformation[J]. Tumour Biol, 2016, 37(9): 12387. doi: 10.1007/s13277-016-5100-4 [6] 张战锋, 谢在春, 庞志宇, 等. NF2基因在丙肝病毒非结构蛋白4B调节细胞周期中的作用[J]. 基础医学与临床, 2019, 39(9): 1289. doi: 10.3969/j.issn.1001-6325.2019.09.014 [7] JANSE VRH, YANG X. The Hippo pathway and cancer immunity: friend or foe?[J]. Oncoscience, 2018, 5(3/4): 49. [8] TAHA Z, JANSE VRH, YANG X. The Hippo pathway: immunity and cancer[J]. Cancers(Basel), 2018, 10(4): 94. [9] HONG AW, MENG Z, PLOUFFE SW, et al. Critical roles of phosphoinositides and NF2 in Hippo pathway regulation[J]. Genes Dev, 2020, 34(7/8): 511. [10] MENG Z, MOROISHI T, GUAN KL. Mechanisms of Hippo pathway regulation[J]. Genes Dev, 2016, 30(1): 1. doi: 10.1101/gad.274027.115 [11] PETRILLI AM, FUSE MA, DONNAN MS, et al. A chemical biology approach identified PI3K as a potential therapeutic target for neurofibromatosis type 2[J]. Am J Transl Res, 2014, 6(5): 471. [12] GUAN Y, GONG Z, XIAO T, et al. Knockdown of miR-572 suppresses cell proliferation and promotes apoptosis in renal cell carcinoma cells by targeting the NF2/Hippo signaling pathway[J]. Int J Clin Exp Pathol, 2018, 11(12): 5705. [13] MIA MM, CHELAKKOT-GOVINDALAYATHIL AL, SINGH MK. Targeting NF2-Hippo/Yap signaling pathway for cardioprotection after ischemia/reperfusion injury[J]. Ann Transl Med, 2016, 4(24): 545. doi: 10.21037/atm.2016.11.85 [14] PETRILLI AM, GARCIA J, BOTT M, et al. Ponatinib promotes a G1 cell-cycle arrest of merlin/NF2-deficient human schwann cells[J]. Oncotarget, 2017, 8(19): 31666. doi: 10.18632/oncotarget.15912 [15] BELTRAMI S, KIM R, GORDON J. Neurofibromatosis type 2 protein, NF2: an uncoventional cell cycle regulator[J]. Anticancer Res, 2013, 33(1): 1. -

 点击查看大图
点击查看大图

图(5)表(3)
计量
- 文章访问数: 3525
- HTML全文浏览量: 1877
- PDF下载量: 15
- 被引次数: 0
