

-
胃癌(gastric cancer, GC)是最常见的危害人类健康的恶性肿瘤之一,根据GLOBOCAN 2020报告显示,GC在恶性肿瘤发病率居第五位,病死率居第四位[1]。目前GC病人的治疗以外科手术为主,辅以放化疗提高GC淋巴结清除的疗效[2]。尽管采取各种手段治疗肿瘤,但GC病人的临床疗效未取得有效进展[3]。因此,研究GC细胞增殖、侵袭、迁移、焦亡等具体机制,对寻找新的治疗靶点,继而提高GC的早诊早治具有重要意义。细胞焦亡(pyroptosis)是一种促炎性程序性细胞死亡(programmed cell death,PCD)[4]。通过激活半胱氨酸天冬蛋白酶-1(caspase-1)信号通路,促进炎症因子释放,引发细胞焦亡[5]。焦亡可能在肿瘤发生及诊疗机制中起着双刃剑的作用[6]。因此,细胞焦亡有望成为治疗肿瘤的思路之一。
炎性小体是细胞焦亡的关键物质,其中NLRP3炎性小体的研究最广泛[7]。NLRP3炎性小体属于模式识别受体NOD样受体(NOD-like receptors,NLRs) 家族成员[8]。与大多数NLRs相似,NLRP3炎性小体作为固有免疫的重要组分在机体免疫反应和疾病发展中发挥着重要作用[9]。目前在多种肿瘤细胞中均可发现NLRP3炎性小体的表达,且其相关蛋白在不同的肿瘤细胞中表现出促癌和抑癌的双重作用[10]。本研究通过检测NLRP3在GC细胞中的表达以及对GC细胞增殖、侵袭、迁移的影响,初步探讨NLRP3炎性小体介导caspase-1依赖的焦亡信号通路在GC的作用机制,以期为GC防治提供理论依据。
-
人GC细胞株MKN45和MGC803以及人胃黏膜上皮细胞株GES-1(中国医学科学院,北京);RPMI 1640培养基和胎牛血清(Gibco, 美国); FAM-FLICA Caspase-1试剂盒(ImmunoChemistry Technologies,美国); 乳酸脱氢酶(LDH)试剂盒(建成生物科技有限公司,中国); NLRP3、caspase-1、GSDMD、白细胞介素(IL)-1β、IL-18(武汉三鹰,中国);siRNA(吉玛制药技术有限公司, 上海);ELISA试剂盒(武汉华美,中国);RIPA细胞裂解液、BCA蛋白定量试剂盒、辣根过氧化物酶标记的二抗、Trizol、CCK8试剂、细胞周期与凋亡试剂盒、结晶紫染液(上海碧云天生物技术有限公司,中国);Lipofectamine2000(Invitrogen,美国);反转录试剂盒(全式金,北京);Transwell小室(康宁,美国);qRT-PCR仪(Roche LightCycler96,Switzerland);流式细胞仪(CYTEK,美国);荧光倒置显微镜(Invitrogen EVOS M5000,美国)。
-
MKN45、MGC803和GES-1细胞用RPMI 1640培养基(含10%胎牛血清-青/链霉素双抗溶液)37℃、5%CO2、97%湿度的培养箱中培养。每2~3 d更换一次培养基,取对数生长期的细胞进行实验。参照Lipo fectamine 2000转染试剂说明书将siNC或siNLRP3转染至MKN45、MGC803细胞。
-
取对数生长期细胞, 以0.5×104个/孔铺于96孔板中。贴壁后转染siRNA,在37 ℃、5% CO2的培养箱中培养24、48、72 h后,根据CCK-8试剂说明书检测450 nm波长处各孔的吸光度(OD)值。
-
收集转染48 h的细胞,上室每孔接种5×104个细胞,加入200μL无血清培养基,下室加入500 μL完全培养基(含10%胎牛血清)。培养24 h后,弃去培养基,PBS缓冲液清洗1次,4%多聚甲醛室温固定20 min,再用湿棉签擦拭去小室内的细胞,然后用0.5%的结晶紫室温染色30 min,PBS洗涤细胞3次之后,晾干,于倒置显微镜下观察并拍照,Image-J计数各组细胞数。
-
将MKN45、MGC803传代至6孔板,待密度达60%~80%转染siRNA。使用胰酶消化、离心收集细胞制备成单细胞悬液,随后参照FAM-FLICA caspase-1分析试剂盒说明,对细胞中活化的caspase-1和碘化丙啶(PI)进行双阳性染色来评估细胞焦亡。
-
将MKN45和MGC803细胞传代至24孔培养板,待密度达60%~80%转染siRNA。按照LDH试剂说明书检测LDH释放,并使用酶标仪检测490 nm OD值。
-
细胞处理结束后,PBS清洗并用胰酶消化,收集细胞,提取总蛋白,BCA法测定蛋白浓度后将蛋白煮沸5 min变性。以50 μg蛋白上样并电泳90 min,再200 mA转膜2 h,用5%脱脂牛奶封闭90 min,TBST清洗3次,每次10 min。加入特异性一抗,4 ℃摇床孵育过夜,TBST清洗3次,加入辣根过氧化物酶标记的二抗,室温孵育2 h,TBST清洗3次,每次10 min。以β-actin为内参对照,ECL曝光显影。
-
Trizol法提取总RNA,逆转录获得cDNA,采用qRT-PCR检测各组细胞NLRP3、caspase-1的mRNA表达,qRT-PCR反应体系和反应程序参照说明书,GAPDH基因作为内参对照,相对表达量按2-△△Ct法计算,引物序列见表 1。
基因 引物序列(5′~3′) NLRP3 正向CGT GAG TCC CAT TAA GAT GGA GT 反向CCC GAC AGT GGA TAT AGA ACA GA caspase-1 正向GAG CAG CCA GAT GGT AGA GC 反向TCT TCA CTT CCT GCC CAC AGA GAPDH 正向CAT GAG AAG TAT GAC AAC AGC CT 反向AGT CCT TCC ACG ATA CCA AAG T 表 1 引物序列
-
收集细胞培养液的上清液,使用人IL-1β、IL-18 ELISA试剂盒进行检测。具体步骤依据试剂盒说明书进行。
-
收集并计数各组细胞,按5×105个/孔的细胞密度在六孔板中进行铺板,细胞密度达60%~80%转染。将各组细胞置于无菌恒温细胞培养箱中培养,用200 μL枪头垂直均匀划线,用PBS洗涤漂浮的细胞3次,分别于0、24、48 h时用光学显微镜拍照。
-
收集并计数各组细胞,按5×105/孔的细胞密度在六孔板中进行铺板,细胞密度达60%~80%转染,24 h后胰蛋白酶消化后离心,用预冷的PBS洗涤3次,加入75%乙醇4 ℃固定过夜,再次使用PBS溶液洗涤3次后按照试剂盒说明操作,避光孵育30 min,最后使用流式细胞仪检测细胞周期分布。
-
采用t检验、方差分析和q检验。
-
为了研究NLRP3炎性小体是否影响GC,本研究首先搜索Oncomine数据库,发现NLRP3在GC中表达上调(P < 0.01)(见表 2)。与GES-1表达情况比较,GC细胞MKN45、MGC803中NLRP3、caspase-1 mRNA和蛋白表达增加,差异均有统计学意义(P < 0.01)(见表 3、图 1)。
分组 n 表达量 正常组 19 0.73±0.01 GC组 71 0.74±0.01 t — 3.87 P — < 0.01 表 2 OncoMine数据库GC中NLRP3表达情况(x±s)
分组 n NLRP3 mRNA caspase-1 mRNA GES-1 3 1.01±0.2 1.01±0.1 MKN45 3 5.05±0.9** 54.75±3.0** MGC803 3 10.11±1.3**## 93.06±1.7**## F — 77.42 1627.29 P — < 0.01 < 0.01 MS组内 — 0.805 3.943 q检验:与GES-1组比较**P < 0.01;与MKN45组比较△△P < 0.01 表 3 GES-1、MKN45、MGC803细胞中NLRP3、caspase-1 mRNA表达情况(x±s)

图 1 Western blotting检测GES-1和MKN45、MGC803细胞NLRP3、caspase-1蛋白表达情况
-
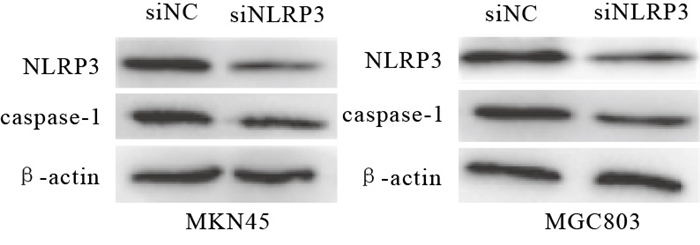
为了探讨NLRP3炎性小体在GC进展中的功能意义,首先采用RNA干扰技术下调NLRP3, 转染至GC细胞株MKN45、MGC803细胞。结果显示,与siNC组相比,siNLRP3组转染后NLRP3、caspase-1 mRNA表达水平均下降,差异均有统计学意义(P < 0.01)(见表 4),NLRP3、caspase-1蛋白表达亦下调(见图 2)。
分组 n NLRP3相对表达量 caspase-1相对表达量 MKN45 MGC803 MKN45 MGC803 siNC组 3 1.00±0.05 1.00±0.03 1.00±0.1 1.02±0.2 siNLRP3组 3 0.36±0.05 0.44±0.03 0.15±0.04 0.26±0.03 t — 15.05 24.14 13.17 6.03 P — < 0.01 < 0.01 < 0.01 < 0.01 表 4 转染NLRP3后MKN45、MGC803细胞中NLRP3、caspase-1 mRNA表达变化(x±s)

图 2 Western blotting检测转染后MKN 45、MGC 803细胞NLRP3、caspase-1蛋白表达情况
-
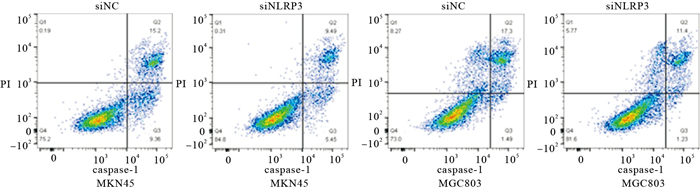
结果显示,与siNC组相比,siNLRP3组MKN45和MGC803的LDH释放减少,且焦亡率降低,差异均有统计学意义(P < 0.01)(见表 5、图 3)。
分组 n LDH释放 焦亡率/% MKN45 MGC803 MKN45 MGC803 siNC组 3 0.71±0.01 0.94±0.01 14.30±0.9 16.87±0.8 siNLRP3组 3 0.54±0.01 0.74±0.01 9.46±0.6 11.17±1.0 t — 18.03 42.43 7.91 7.69 P — < 0.01 < 0.01 < 0.01 < 0.01 表 5 转染后MKN45、MGC803细胞LDH释放OD值及caspase-1/PI双染阳性细胞比例的情况(x±s)

图 3 流式细胞术检测下调NLRP3对GC细胞焦亡影响(caspase-1/PI染色)
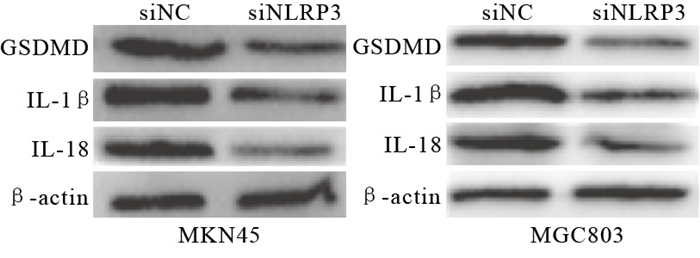
进一步对焦亡相关蛋白进行检测,与siNC组相比,siNLRP3组GSDMD、IL-1β、IL-18表达下降(见图 4)。对培养基上清液中细胞因子IL-1β、IL-18水平进行检测,结果显示siNLRP3组表达下降(P < 0.01)(见表 6)。

图 4 Western blotting检测GSDMD、IL-1β、IL-18蛋白表达情况
分组 n IL-1β IL-18 MKN45 MGC803 MKN45 MGC803 siNC组 3 6 504.0±115.21 6 673.28±31.53 95.35±1.35 127.02±1.68 siNLRP3组 3 5 408.0±35.09 5 448.35±10.61 63.88±1.08 78.67±1.37 t — 15.76 63.77 31.47 38.71 P — < 0.01 < 0.01 < 0.01 < 0.01 表 6 转染后MKN45、MGC803细胞培养基上清液IL-1β、IL-18水平情况(x±s;pg/mL)
-
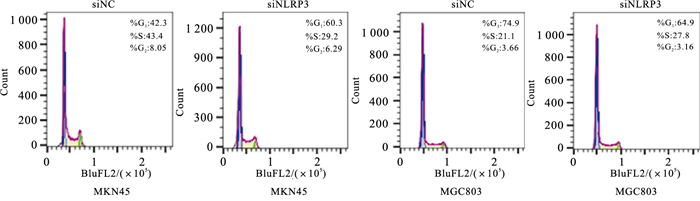
转染后,siNC组和siNLRP3组MKN45、MGC803的OD值均随时间延长而增加,且与siNC组比较,相同作用时间下siNLRP3组GC细胞OD值降低,即细胞活力降低,差异均有统计学意义(P < 0.01)(见表 7);细胞周期实验显示阻滞在G1期(P < 0.01)(见图 5、表 8)。与siNC组相比,siNLRP3组MKN45、MGC803迁移率降低,穿过小室的GC细胞数减少,差异有统计学意义(P < 0.05~P < 0.01)(见表 9)。
分组 n 0 h 24 h 48 h 72 h F P MS组内 MKN45 siNC组 3 0.20±0.01 0.80±0.01** 1.23±0.04**## 1.83±0.03**##△△ 2 137.88 < 0.01 0.001 siNLRP3组 3 0.20±0.01 0.45±0.01** 0.68±0.03**## 1.07±0.09**##△△ 204.60 < 0.01 0.002 t — — 53.00 18.81 14.84 — — — P — — < 0.01 < 0.01 < 0.01 — — — MGC803 siNC组 3 0.20±0.01 0.75±0.03** 1.34±0.03**## 2.03±0.06**##△△ 1 322.15 < 0.01 0.001 siNLRP3组 3 0.21±0.01 0.52±0.01** 0.80±0.03**## 1.46±0.01**##△△ 2 189.01 < 0.01 0.001 t — — 12.00 19.09 17.02 — — — P — — < 0.01 < 0.01 < 0.01 — — — q检验:与0 h比较**P < 0.01;与24 h比较##P < 0.01;与48 h比较△△P < 0.01 表 7 转染后MKN45、MGC803细胞增殖情况OD值(x±s)

图 5 流式细胞术检测转染后细胞周期阻滞情况
分组 n MKN45细胞G1/% MGC803细胞G1/% siNC组 3 42.30±0.4 76.53±2.10 siNLRP3组 3 59.57±1.81 64.87±0.55 t — 16.09 9.31 P — < 0.01 < 0.01 表 8 siRNLRP3对MKN45、MGC803细胞周期G1期的影响(x±s)
分组 n MKN45迁移率/% MGC803迁移率/% MKN45 MGC803 24 h 48 h 24 h 48 h 细胞数 细胞数 siNC组 3 21.60±3.81 29.93±1.31 29.87±2.52 38.21±1.52 307.67±27.30 390.67±36.56 siNLRP3组 3 8.93±1.62 22.97±0.55 17.03±4.69 32.41±2.68 150.67±14.47 137.67±10.02 t — 5.30 8.51 4.18 3.26 8.80 11.56 P — < 0.01 < 0.01 < 0.05 < 0.05 < 0.01 < 0.01 表 9 siNLRP3对MKN45、MGC803细胞迁移、侵袭能力的影响(x±s)
-
GC的发生发展与多种因素(如幽门螺旋杆菌、吸烟、饮酒等)相关[11-12]。慢性炎症可能是GC发生的主要原因之一,组织细胞长期暴露在炎症环境中,会增加癌症的风险[13]。在慢性炎症反应中,大量炎症因子从炎症细胞释放,并通过激活各种细胞信号传导途径引起细胞生存微环境发生变化,调节细胞代谢过程,进一步来促进肿瘤细胞的增殖和侵袭迁移[14]。
近年来,细胞焦亡引起了广泛关注,细胞焦亡在肿瘤的发生发展的不同阶段起着不可或缺的作用[15]。细胞焦亡主要包括依赖caspase-1的经典信号通路以及依赖caspase-4/5/11的非经典信号通路[4]。当发生焦亡后,细胞膜丧失完整性,出现孔洞,细胞发生肿胀和破裂,细胞内物质被释放到细胞外,引起炎症反应[16]。NLRP3炎性小体是细胞焦亡过程的关键物质,是由NLRP3、凋亡相关斑点样蛋白(apoptosis-associated speck-like protein containing a card, ASC)和pro-caspase-1组成的多蛋白复合物[17],在病原相关分子模式和危险相关分子模式刺激下,NLRP3炎性小体组装,使pro-caspase-1活化,活化的caspase-1一方面可以切割GSDMD,另一方面切割pro-IL-1β、pro-IL-18,形成有活性的IL-1β、IL-18,引起炎症反应以及细胞焦亡[18-19]。NLRP3炎性小体参与多种肿瘤发生发展过程,如肺癌[20]、结直肠癌[21]、乳腺癌[22]等。然而,NLRP3在不同的癌症中起着有益或有害的作用,NLRP3炎性小体通路的激活和IL-1β的分泌可以促进乳腺癌进展和肺转移[7];在肺癌细胞A549中,NLRP3炎性小体的激活可促进A549的增殖和迁移[23];NLRP3炎性小体通过促进自然杀伤细胞的杀瘤活性可抑制结直肠癌在肝脏中的转移生长[10]。NLRP3炎性小体在GC中具体作用机制及其与肿瘤发生发展的关系值得进一步研究。
本研究结果显示,NLRP3、caspase-1在GC细胞中表达增加,下调NLRP3后MKN45、MGC803细胞NLRP3、caspase-1表达受到抑制。同时LDH释放降低,细胞焦亡率降低,焦亡相关蛋白GSDMD、IL-1β、IL-18的表达水平降低。NLRP3下调后GC细胞的增殖、侵袭、迁移能力降低。提示NLRP3可能通过介导caspase-1依赖的细胞焦亡影响GC细胞的增殖、侵袭和迁移。
综上所述,NLRP3表达的上调与GC相关,是GC病人预后不良的分子标志之一。下调NLRP3可以抑制GC细胞焦亡和炎症反应,与此同时增殖、侵袭及迁移能力随之降低。本研究为基于NLRP3炎性小体治疗GC提供了参考,还需要更多研究来更好地理解NLRP3/caspase-1介导的焦亡通路表达。
NLRP3炎性小体在胃癌细胞中的表达及对其焦亡、增殖、侵袭、迁移的影响
Expression of NLRP3 inflammasome in gastric cancer cells and its effect on the pyrophosis, proliferation, invasion and migration
-
摘要:
目的 研究胃癌细胞中NLRP3炎性小体的表达及其对增殖、侵袭和迁移的影响。并初步探讨NLRP3介导caspase-1依赖的焦亡信号通路在胃癌发展中的作用。 方法qRT-PCR和Western blotting法分别检测人胃黏膜上皮细胞(GES-1)和人胃癌细胞(MKN45及MGC803)中NLRP3、caspase-1表达;随后下调NLRP3,检测胃癌细胞NLRP3、caspase-1 mRNA和蛋白的表达;Western blotting检测GSDMD、白细胞介素(IL)-18、IL-1β蛋白表达;ELISA法检测细胞培养液上清液中IL-1β、IL-18含量;乳酸脱氢酶(LDH)实验、流式细胞术检测细胞焦亡;CCK-8法及细胞周期试剂盒检测细胞增殖情况;Transwell和划痕实验检测胃癌细胞侵袭和迁移能力。 结果NLRP3、caspase-1在胃癌细胞中较GES-1表达增高(P < 0.01)。下调NLRP3后,胃癌细胞LDH释放率及焦亡率显著降低(P < 0.01);GSDMD、IL-1β、IL-18表达降低(P < 0.01);细胞增殖阻滞在G1期(P < 0.01);细胞增殖以及侵袭迁移能力显著减弱。 结论NLRP3在胃癌中高表达,其表达影响胃癌细胞的增殖、侵袭和迁移以及其介导的caspase-1依赖焦亡信号通路关键蛋白的表达。 Abstract:ObjectiveTo investigate the expression of NLRP3 inflammasome in gastric cancer(GC) and its effects on the proliferation, invasion and migration of cells, and the role of NLRP3-mediated caspase-1-dependent pyroptosis signaling pathway in the development of GC. MethodsThe expression levels of NLRP3 and caspase-1 in human gastric epithelial cells(GES-1) and human gastric cancer cells(MKN45 and MGC803) were detected by qRT-PCR and Western blotting.After the expression level of NLRP3 was down-regulated, and the mRNA and protein expression levels of NLRP3 and caspase-1 were detected, the protein expression levels of GSDMD, IL-18 and IL-1β were detected using Western blotting.The content of IL-1β and IL-18 in supernatant were detected by ELISA, the pyroptosis was detected by LDH and flow cytometry, and the cell proliferation was detected by CCK-8 assay and cell cycle kit.Transwell and wound healing were used to detect the invasion and migration of cells. ResultsThe expression levels of NLRP3 and caspase-1 in GC cells were higher than those in GES-1(P < 0.01).After the expression level of NLRP3 was downregulated, the release LDH rate and pyroptosis rate of LDH significantly decreased(P < 0.01), and the expression levels of GSDMD, IL-1β and IL-18 decreased(P < 0.01).Compared with the NC group, the cell proliferation was arrested in G1 phase(P < 0.01).The proliferation, invasion and migration of siNLRP3 cells were significantly reduced. ConclusionsThe expression of NLRP3 in GC cells is high, and its expression affects the proliferation, invasion and migration of GC cells and expression of key proteins in caspase-1-dependent pyroptosis signaling pathway mediated by NLRP3. -
Key words:
- gastric neoplasms /
- NLRP3 inflammasome /
- pyroptosis /
- caspase-1
-
表 1 引物序列
基因 引物序列(5′~3′) NLRP3 正向CGT GAG TCC CAT TAA GAT GGA GT 反向CCC GAC AGT GGA TAT AGA ACA GA caspase-1 正向GAG CAG CCA GAT GGT AGA GC 反向TCT TCA CTT CCT GCC CAC AGA GAPDH 正向CAT GAG AAG TAT GAC AAC AGC CT 反向AGT CCT TCC ACG ATA CCA AAG T  下载: 导出CSV
下载: 导出CSV
表 2 OncoMine数据库GC中NLRP3表达情况(x±s)
分组 n 表达量 正常组 19 0.73±0.01 GC组 71 0.74±0.01 t — 3.87 P — < 0.01
下载: 导出CSV
表 3 GES-1、MKN45、MGC803细胞中NLRP3、caspase-1 mRNA表达情况(x±s)
分组 n NLRP3 mRNA caspase-1 mRNA GES-1 3 1.01±0.2 1.01±0.1 MKN45 3 5.05±0.9** 54.75±3.0** MGC803 3 10.11±1.3**## 93.06±1.7**## F — 77.42 1627.29 P — < 0.01 < 0.01 MS组内 — 0.805 3.943 q检验:与GES-1组比较**P < 0.01;与MKN45组比较△△P < 0.01
下载: 导出CSV
表 4 转染NLRP3后MKN45、MGC803细胞中NLRP3、caspase-1 mRNA表达变化(x±s)
分组 n NLRP3相对表达量 caspase-1相对表达量 MKN45 MGC803 MKN45 MGC803 siNC组 3 1.00±0.05 1.00±0.03 1.00±0.1 1.02±0.2 siNLRP3组 3 0.36±0.05 0.44±0.03 0.15±0.04 0.26±0.03 t — 15.05 24.14 13.17 6.03 P — < 0.01 < 0.01 < 0.01 < 0.01
下载: 导出CSV
表 5 转染后MKN45、MGC803细胞LDH释放OD值及caspase-1/PI双染阳性细胞比例的情况(x±s)
分组 n LDH释放 焦亡率/% MKN45 MGC803 MKN45 MGC803 siNC组 3 0.71±0.01 0.94±0.01 14.30±0.9 16.87±0.8 siNLRP3组 3 0.54±0.01 0.74±0.01 9.46±0.6 11.17±1.0 t — 18.03 42.43 7.91 7.69 P — < 0.01 < 0.01 < 0.01 < 0.01
下载: 导出CSV
表 6 转染后MKN45、MGC803细胞培养基上清液IL-1β、IL-18水平情况(x±s;pg/mL)
分组 n IL-1β IL-18 MKN45 MGC803 MKN45 MGC803 siNC组 3 6 504.0±115.21 6 673.28±31.53 95.35±1.35 127.02±1.68 siNLRP3组 3 5 408.0±35.09 5 448.35±10.61 63.88±1.08 78.67±1.37 t — 15.76 63.77 31.47 38.71 P — < 0.01 < 0.01 < 0.01 < 0.01
下载: 导出CSV
表 7 转染后MKN45、MGC803细胞增殖情况OD值(x±s)
分组 n 0 h 24 h 48 h 72 h F P MS组内 MKN45 siNC组 3 0.20±0.01 0.80±0.01** 1.23±0.04**## 1.83±0.03**##△△ 2 137.88 < 0.01 0.001 siNLRP3组 3 0.20±0.01 0.45±0.01** 0.68±0.03**## 1.07±0.09**##△△ 204.60 < 0.01 0.002 t — — 53.00 18.81 14.84 — — — P — — < 0.01 < 0.01 < 0.01 — — — MGC803 siNC组 3 0.20±0.01 0.75±0.03** 1.34±0.03**## 2.03±0.06**##△△ 1 322.15 < 0.01 0.001 siNLRP3组 3 0.21±0.01 0.52±0.01** 0.80±0.03**## 1.46±0.01**##△△ 2 189.01 < 0.01 0.001 t — — 12.00 19.09 17.02 — — — P — — < 0.01 < 0.01 < 0.01 — — — q检验:与0 h比较**P < 0.01;与24 h比较##P < 0.01;与48 h比较△△P < 0.01
下载: 导出CSV
表 8 siRNLRP3对MKN45、MGC803细胞周期G1期的影响(x±s)
分组 n MKN45细胞G1/% MGC803细胞G1/% siNC组 3 42.30±0.4 76.53±2.10 siNLRP3组 3 59.57±1.81 64.87±0.55 t — 16.09 9.31 P — < 0.01 < 0.01
下载: 导出CSV
表 9 siNLRP3对MKN45、MGC803细胞迁移、侵袭能力的影响(x±s)
分组 n MKN45迁移率/% MGC803迁移率/% MKN45 MGC803 24 h 48 h 24 h 48 h 细胞数 细胞数 siNC组 3 21.60±3.81 29.93±1.31 29.87±2.52 38.21±1.52 307.67±27.30 390.67±36.56 siNLRP3组 3 8.93±1.62 22.97±0.55 17.03±4.69 32.41±2.68 150.67±14.47 137.67±10.02 t — 5.30 8.51 4.18 3.26 8.80 11.56 P — < 0.01 < 0.01 < 0.05 < 0.05 < 0.01 < 0.01
下载: 导出CSV
-
[1] SUNG H, FERLAY J, SIEGEL RL, et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209. doi: 10.3322/caac.21660 [2] DIGKLIA A, WAGNER AD. Advanced gastric cancer: Current treatment landscape and future perspectives[J]. World J Gastroenterol, 2016, 22(8): 2403. doi: 10.3748/wjg.v22.i8.2403 [3] CREW KD, NEUGUT AL. Epidemiology of upper gastrointestinal malignancies[J]. Semin Oncol, 2004, 31(4): 450. doi: 10.1053/j.seminoncol.2004.04.021 [4] WANG Y, GAO W, SHI X, et al. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin[J]. Nature, 2017, 547(7661): 99. doi: 10.1038/nature22393 [5] KOPALLI SR, KANG TB, LEE KH, et al. NLRP3 inflammasome activation inhibitors in inflammation-associated cancer immunotherapy: an update on the recent patents[J]. Recent Pat Anticancer Drug Discov, 2018, 13(1): 106. [6] HUMPHRIES F, SHMUEL-GALIA L, KETELUT-CARNEIRO N, et al. Succination inactivates gasdermin D and blocks pyroptosis[J]. Science, 2020, 369(6511): 1633. doi: 10.1126/science.abb9818 [7] ERSHAID N, SHARON Y, DORON H, et al. NLRP3 inflammasome in fibroblasts links tissue damage with inflammation in breast cancer progression and metastasis[J]. Nat Commun, 2019, 10(1): 4375. doi: 10.1038/s41467-019-12370-8 [8] SHARIF H, WANG L, WANG WL, et al. Structural mechanism for NEK7-licensed activation of NLRP3 inflammasome[J]. Nature, 2019, 570(7761): 338. doi: 10.1038/s41586-019-1295-z [9] REN G, ZHANG X, XIAO Y, et al. ABRO1 promotes NLRP3 inflammasome activation through regulation of NLRP3 deubiquitination[J]. EMBO J, 2019, 38(6): e100371. [10] DUPAUL-CHICOINE J, ARABZADEH A, DAGENAIS M, et al. The Nlrp3 inflammasome suppresses colorectal cancer metastatic growth in the Liver by promoting natural killer cell tumoricidal activity[J]. Immunity, 2015, 43(4): 751. doi: 10.1016/j.immuni.2015.08.013 [11] LI S, LIANG X, MA L, et al. MiR-22 sustains NLRP3 expression and attenuates H. pylori-induced gastric carcinogenesis[J]. Oncogene, 2018, 37(7): 884. doi: 10.1038/onc.2017.381 [12] 任志, 马振增, 王启之. 幽门螺杆菌感染病人胃癌组织中MMP-7、VEGF的表达及相关性研究[J]. 蚌埠医学院学报, 2020, 45(6): 739. [13] WAGNER EF, NEBREDA ÁR. Signal integration by JNK and p38 MAPK pathways in cancer development[J]. Nat Rev Cancer, 2009, 9(8): 537. doi: 10.1038/nrc2694 [14] LIM W, AN Y, YANG C, et al. Chrysophanol induces cell death and inhibits invasiveness via mitochondrial calcium overload in ovarian cancer cells[J]. J Cell Biochem, 2018, 119(12): 10216. doi: 10.1002/jcb.27363 [15] 王石健, 徐蒙娜, 汪佳兵. gasdermin D和gasdermin E介导的细胞焦亡在肿瘤中的研究进展[J]. 中国临床药理学与治疗学. 2020, 25(3): 352. [16] KARKI R, MAN SM, KANNEGANTI TD. Inflammasomes and Cancer[J]. Cancer Immunol Res, 2017, 5(2): 94. doi: 10.1158/2326-6066.CIR-16-0269 [17] 康品方, 戎李, 孙硕等. 血清ALDH2和NLRP3活性水平在冠心病不同血管病变程度中的变化[J]. 蚌埠医学院学报. 2020, 45(7): 854. [18] HE Y, ZENG MY, YANG D, et al. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux[J]. Nature, 2016, 530(7590): 354. doi: 10.1038/nature16959 [19] TENG JF, MEI QB, ZHOU XG, et al. Polyphyllin Ⅵ induces caspase-1-mediated pyroptosis via the induction of ROS/NF-κB/NLRP3/GSDMD signal axis in non-small cell lung cancer[J]. Cancers(Basel), 2020, 12(1): 193. [20] ZHANG CC, LI CG, WANG YF, et al. Chemotherapeutic paclitaxel and cisplatin differentially induce pyroptosis in A549 lung cancer cells via caspase-3/GSDME activation[J]. Apoptosis, 2019, 24(3/4): 312. [21] YU J, LI S, QI J, et al. Cleavage of GSDME by caspase-3 determines lobaplatin-induced pyroptosis in colon cancer cells[J]. Cell Death Dis, 2019, 10(3): 193. doi: 10.1038/s41419-019-1441-4 [22] WU X, MAO X, HUANG Y, et al. Detection of proteins associated with the pyroptosis signaling pathway in breast cancer tissues and their significance[J]. Int J Clin Exp Pathol, 2020, 13(6): 1408. [23] WANG Y, KONG H, ZENG X, et al. Activation of NLRP3 inflammasome enhances the proliferation and migration of A549 lung cancer cells[J]. Oncol Rep, 2016, 35(4): 2053. doi: 10.3892/or.2016.4569 -

 点击查看大图
点击查看大图

图(5)表(9)
计量
- 文章访问数: 4502
- HTML全文浏览量: 2570
- PDF下载量: 15
- 被引次数: 0
