



-
胃癌是消化系统最常见的恶性肿瘤,是全球第三大癌症死亡原因[1]。我国癌症报告显示,胃癌位居我们恶性肿瘤发病率和死亡率的第2位,仅次于肺癌,造成了巨大的经济以及公共卫生负担[2]。Periostin(POSTN)为成骨细胞特异性因子2[3]。研究[4]发现POSTN参与细胞黏附、结缔组织的发育、细胞存活、血管形成、组织修复和再生。且POSTN在乳腺癌、肺癌、结肠癌、胰腺癌和卵巢癌等癌症中过表达[5]。上皮间质转化(EMT)在癌细胞的增殖、浸润和远处转移过程中发挥重要作用[6],以及蛋白激酶B(AKT)信号通路在胃癌的癌变和细胞生存控制过程中的发挥着重要作用[7]。本研究探讨了POSTN调控AKT信号通路激活胃癌细胞EMT的机制。现作报道。
-
人胃组织收集于蚌埠医学院第一附属医院普外科,并已获得病人的知情同意。人胃癌细胞MGC 803(购自北京协和细胞库)、RPMI-1640培养基和胎牛血清(美国Gibco公司)、过表达和敲除POSTN基因的慢病毒(吉凯基因)、Trizol(美国Invitrogen公司)、PCR引物(上海生工生物工程股份有限公司)、SYBR Green PCR Master Mix和RevertAid First Strand cDNA Synthesis Kit试剂盒(北京天根生化科技有限公司)、MTT试剂和结晶紫染料(北京索莱宝生物科技有限公司)、Transwell小室(美国corning公司)。
-
将胃癌组织置于4%多聚甲醛固定、脱水机脱水、石蜡包埋、制片、烤片和抗原修复后,孵育POSTN(stanta cruz biotechnology,1∶100)和CD44(abcam,1∶200),洗涤3次后,孵育Alexa FluorⓇ 594标记兔抗山羊IgG(H+L)和FITC标记山羊抗小鼠IgG(H+L)(北京中杉金桥生物有限公司,1∶200),洗涤后滴加DAPI(thermo)对细胞核进行染色,置于激光共聚焦显微镜(Leica,SP8)采集图片。
-
将MGC 803细胞移入含有10%胎牛血清的RPMI-1640培养基中,置于含有5% CO2、37 ℃的培养箱中培养。
-
将MGC 803细胞种植于6孔板中,提前1 h换新鲜的培养,分别滴加过表达和敲除POSTN基因的慢病毒溶液,置于培养箱中培养6 h,更换新鲜的培养基。培养3 d后,更换含有5 μg/mL嘌呤霉素的完全培养基进行阳性细胞筛选,收集各组细胞进行后续实验。
-
将细胞种植6孔板中,更换新鲜的无血清RPMI-1640培养基置于培养箱中培养24 h,更换含有LY294002(sigma,10 μmol/L)的无血清RPMI-1640培养基于培养箱中培养24 h,用于后续实验。
-
采用Trizol法提取MGC 803、过表达POSTN基因的MGC 803和敲除POSTN基因的MGC 803以及添加LY294002抑制剂的胃癌细胞,按照PCR说明书进行PCR实验。GAPDH作为内参,所用的引物序列见表 1。
基因名称 上游序列 下游序列 GAPDH CTG GGC TAC ACT GAG CAC C AAG TGG TCG TTG AGG GCA ATG CD44 CTG CCG CTT TGC AGG TGT A CAT TGT GGG CAA GGT GCT ATT POSTN CTC ATA GTC GTA TCA GGG GTC G ACA CAG TCG TTT TCT GTC CAC α-sma CCT GTG TTG TGG TTT ACA CTG G GGG GGA ATT ATC TTT CCT GGT CC VIM AGT CCA CTG AGT ACC GGA GAC CAT TTC ACG CAT CTG GCG TTC Snail TCG GAA GCC TAA CTA CAG CGA AGA TGA GCA TTG GCA GCG AG Slug CGA ACT GGA CAC ACA TAC AGT G CTG AGG ATC TCT GGT TGT GGT CDH1 ATT TTT CCC TCG ACA CCC GAT TCC CAG GCG TAG ACC AAG A 表 1 引物序列
-
采用RAPI裂解液提取胃癌细胞,经BCA蛋白定量、loading buffer稀释和煮沸变性后经SDS-PAGE电泳与转膜后,孵育POSTN(1∶1 000)、CD44(1∶1 000)、α-sma(CST,1∶1 000)、VIM(CST,1∶1 000)、snail+slug(CST,1∶1 000)、E-cad(CST,1∶1 000)和GAPDH(CST,1∶1 000),在经过氧化物酶连接的二抗(北京中杉金桥生物有限公司,1∶2 000)孵育,经暗室曝光并采集图片。
-
将胃癌细胞种植于96孔板中,每孔1×103个细胞,置于培养箱中培养24 h,每孔加入10 μL的CCK-8试剂,培养箱中进行培养4 h,在490 nm下经酶标仪检测吸光度(OD)值。
-
消化稀释胃癌细胞,用无血清的RPMI-1640培养基重悬细胞,取4×104/mL的细胞100 μL种植于Transwell小室中,在24孔板中加入1 mL的完全培养,并种植好细胞的Transwell小室置于24孔板中,培养24 h后,再经多聚甲醛固定和结晶紫染色后,再显微镜下随机选取5个视野采集图片并统计分析。侵袭: 冰上完全溶解Matrigel基质胶,取40 μL加入预冷的Transwell小室中;其余的操作步骤同迁移实验。
-
采用方差分析和q检验。
-
与癌旁组织相比,原发灶和转移灶组织中POSTN的表达均明显地升高,以及转移灶组织中POSTN表达也明显高于原发灶组织,且POSTN的表达与CD44(肿瘤干性标志物)有共定位现象(见图 1)。

图 1 免疫荧光显示POSTN和CD44在胃癌组织中的表达
-
免疫荧光显示胃癌原发灶组织临床分期越晚,POSTN的表达越高,且POSTN的表达与CD44也有共定位现象(见图 2)。

图 2 免疫荧光显示POSTN和CD44在不同临床分期的胃癌组织中的表达
-
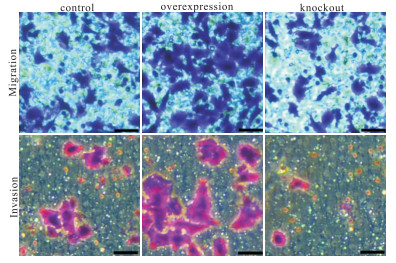
与对照组相比,过表达POSTN基因MGC 803的细胞增殖活性、迁移和侵袭能力明显地升高;敲除POSTN基因MGC 803的细胞增殖活性、迁移和侵袭能力细胞明显地降低(P < 0.01)(见图 3和表 2~4)。

图 3 POSTN促进MGC 803细胞迁移和侵袭
分组 n OD值 对照组 3 1.06±0.21 过表达组 3 1.80±0.24** 敲除组 3 0.68 ±0.18**△△ F — 25.92 P — < 0.01 MS组内 — 0.045 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 2 CCK8检测调控POSTN基因对MGC 803细胞增殖的影响(x±s)
分组 n 细胞个数 对照组 3 557.33±125.10 过表达组 3 1 004.67±143.14** 敲除组 3 328.67±94.88**△△ F — 24.67 P — < 0.05 MS组内 — 14 448.000 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 3 调控POSTN基因对MGC 803细胞转移的影响(x±s)
分组 n 细胞个数 对照组 3 110.00±44.00 过表达组 3 550.00±166.10** 敲除组 3 55.33±11.02**△△ F — 22.33 P — < 0.05 MS组内 — 9 981.778 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 4 调控POSTN基因对MGC 803细胞侵袭的影响
-
与对照组相比,过表达POSTN基因MGC 803的间质标志物(α-sma、VIM、snail和slug)的表达升高,上皮标注物E-Cadherin表达下降;而敲除POSTN基因MGC 803的间质标志物(α-sma、VIM、snail和slug)的表达下降,上皮标注物E-Cadherin表达升高(P < 0.01)(见表 5、图 4)。
分组 n POSTN CD44 α-SMA VIM Snail Slug CDH1 对照组 3 1 1 1 1 1 1 1 敲除组 3 0.01±0.01** 0.58±0.32** 0.57±0.17** 0.48±0.15** 0.66±0.10** 0.57±0.31** 1.96±0.38** 过表达组 3 7.66±1.95**△△ 3.30±1.00**△△ 2.88±0.57**△△ 2.67±0.47**△△ 1.88±0.24**△△ 2.10±0.32**△△ 0.55±0.08**△△ F — 41.01 17.48 38.62 49.30 53.48 28.20 31.06 P — < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 1.268 0.369 0.117 0.080 0.022 0.066 0.050 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 5 RT-qPCR检测POSTN对MGC 803细胞EMT相关基因的影响(x±s)

图 4 POSTN促进MGC 803细胞EMT
-
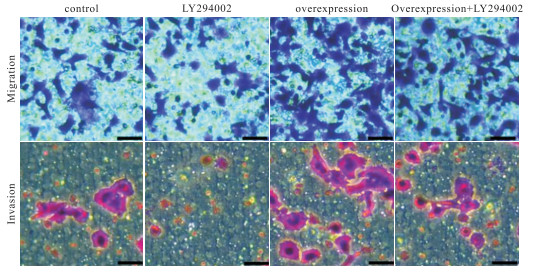
分别与正常的和过表达POSTN基因的MGC 803相比,添加LY294002会明显地降低细胞的增殖、转移和侵袭能力(P < 0.01)(见图 5、表 6)。

图 5 LY294002抑制POSTN激活MGC 803的细胞增殖、迁移和侵袭
分组 n OD值 对照组 3 1.28±1.28 LY294002 3 0.61±0.15** 过表达组 3 2.34±0.17** 过表达组+LY294002 3 1.51±0.17**△△ F — 60.23 P — < 0.01 MS组内 — 0.028 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 6 CCK8检测LY294002对MGC 803细胞调控POSTN基因后增殖的影响(x±s)
-
Western blotting和RT-qPCR显示,分别与正常的和过表达POSTN基因的MGC 803相比,添加LY294002会明显地降低间质标志物(α-sma、VIM、snail和slug)的表达,以及升高上皮标志物E-Cadherin的表达(P < 0.01)(见图 6、表 7~9)。
分组 n 细胞个数 对照组 3 412.33±63.12 LY294002 3 269.67±36.23** 过表达组 3 831.33±87.30** 过表达组+LY294002 3 460.33±116.07△△ F — 26.06 P — < 0.01 MS组内 — 6 597.580 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 7 LY294002对MGC 803细胞的调控POSTN基因后转移的影响(x±s)
分组 n 细胞个数 对照组 3 107.67±24.01 LY294002 3 40.00±19.97** 过表达组 3 270.67±67.00** 过表达组+LY294002 3 136.67±37.22△△ F — 16.42 P — < 0.01 MS组内 — 1 712.500 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 8 LY294002对MGC 803细胞的调控POSTN基因后迁移的影响(x±s)
分组 n POSTN CD44 α-SMA VIM Snail Slug CDH1 AKT 对照组 3 1 1 1 1 1 1 1 1 LY294002 3 1.02±0.18** 0.60±0.16** 0.49±0.15** 0.55±0.11** 0.64±0.07** 0.62±0.10** 1.92±0.64** 0.12±0.08** 过表达组 3 6.45±1.81** 3.34±0.89** 2.92±0.35** 2.82±0.17** 2.14±0.46** 2.17±0.19** 0.52±0.11** 1.03±0.20** 过表达组+LY294002 3 6.30±2.46△△ 1.68±0.39△△ 1.38±0.25△△ 1.22±0.18△△ 1.06±0.09△△ 0.95±0.09△△ 1.11±0.18△△ 0.53±0.11△△ F — 12.32 18.13 64.06 161.55 22.50 101.98 8.89 38.90 P — < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 2.336 0.242 0.051 0.018 0.055 0.014 0.115 0.014 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01 表 9 RT-qPCR检测LY294002抑制POSTN激活MGC 803细胞的EMT(x±s)
-
POSTN已被证实在各种转移性肿瘤的癌症基质中上调,例如肝细胞癌、肝内胆管癌、食道鳞状细胞癌、结肠癌、神经胶质瘤、乳腺癌、肺癌、黑素瘤、胰腺癌、卵巢癌、膀胱癌癌、头颈癌、甲状腺癌和前列腺癌[8]。POSTN被认为在肿瘤的发生、肿瘤细胞浸润性、肿瘤细胞转移性、血管生成、淋巴结血管生成、化疗耐药性和病人复发中均起着重要作用[9]。本研究进行了POSTN和CD44共同免疫荧光染色,以检测POSTN在胃癌转移灶组织、原发灶组织及其邻近正常组织中的表达。与邻近的正常组织相比,POSTN在原发灶和转移灶胃癌组织中的表达更高。对不同临床阶段胃癌病人的原发灶组织中的POSTN蛋白进行了免疫荧光染色,发现晚期胃癌病人的原发灶组织中POSTN的蛋白表达水平明显地高于早期。这些结果表明POSTN可能在胃癌的进展中起重要作用。
为了进一步验证POSTN在胃癌发生中的作用,我们对MGC 803进行过表达和敲除POSTN基因,发现过表达POSTN基因可以增强胃癌细胞的增殖活性,以及促进胃癌细胞的迁移和侵袭;并且敲除POSTN基因会降低胃癌细胞的增殖活性和减弱胃癌细胞的迁移和侵袭。这些实验结果表明POSTN促进了胃癌细胞的增殖、迁移和侵袭,为胃癌提供了支持肿瘤生长以及促进胃癌细胞增殖和转移的微环境。
EMT是指上皮细胞失去根尖-基底极性和细胞间质黏附,转化为具有侵袭性间充质细胞的过程[10]。EMT在肿瘤的进展、转移和耐药性中发挥着重要作用[11]。本研究发现,与正常细胞相比,过表达POSTN会促进胃癌细胞间质标志物的表达和抑制上皮标志物的表达;以及敲除POSTN会抑制胃癌细胞间质标志物的表达和促进上皮标志物的表达。这些结果提示POSTN会促进胃癌细胞的EMT。然而POSTN如何影响EMT的,目前尚不清楚。另有研究[12]表明,AKT信号通路参与胃癌细胞增殖、肿瘤发生和预后,我们发现AKT抑制剂LY2940002明显缓解过表达POSTN导致的胃癌细胞增殖活性、迁移和侵袭能力,以及抑制胃癌细胞间质标志物的表达和促进上皮标志物的表达。这些结果表明POSTN可以经AKT信号通路调控胃癌细胞的EMT,进而促进胃癌细胞的增殖、转移和侵袭。
综上所述,POSTN不仅促进胃癌的转移,还加剧了癌症的行为。最重要的是,POSTN的高表达与晚期胃癌有关,POSTN的高表达水平是胃癌的危险因素之一。因此,POSTN可能是胃癌肿瘤浸润、转移和组织学分化的预测因子。这表明POSTN是恶性肿瘤及其转移的潜在治疗靶标。
Periostin调控AKT信号通路激活胃癌细胞的上皮间质转化
Periostin regulates AKT signaling pathway to activate epithelial-mesenchymal transition of gastric cancer cells
-
摘要:
目的探讨Periostin (POSTN)调控AKT信号通路激活胃癌细胞的上皮间质转化(EMT)的机制。 方法选取胃癌病人的癌旁、原发灶、转移灶以及不同临床分期的原发灶胃癌组织标本, 采用POSTN与CD44共同免疫荧光染色。选择人胃癌细胞MGC 803细胞, 并分别过表达和敲除POSTN基因, 分别添加AKT抑制剂LY294002进行孵育培养, 采用RT-qPCR法检测各组细胞的POSTN、CD44和EMT相关基因的表达水平; 采用蛋白印迹技术检测各组细胞的POSTN、CD44和EMT相关蛋白的表达水平; 采用CCK-8检测各组细胞的增殖水平; 采用Transwell实验检测细胞迁移和侵袭能力。 结果胃癌病人的转移灶中POSTN表达高于原发灶, 且原发灶中POSTN表达高于癌旁组织。与MGC 803细胞相比, 过表达POSTN基因的胃癌细胞中POSTN、CD44、α-sma、snail、slug、Vimentin表达升高和E-Cadherin表达降低以及增殖活性、迁移和侵袭能力升高; 敲除POSTN基因的胃癌细胞中CD44、α-sma、snail、slug、Vimentin表达降低以及E-Cadherin表达升高以及增殖活性、迁移和侵袭能力降低; 添加LY294002会显著地改善胃癌细胞中过表达POSTN基因引起的α-sma、snail、slug、Vimentin、E-Cadherin表达变化和增殖活性及迁移与侵袭能力的改变(P < 0.01)。 结论胃癌原发灶和转移灶中POSTN表达上调且临床分期越晚的原发灶中POSTN表达越高; POSTN通过调控AKT信号通路激活胃癌细胞的EMT, 进而促进胃癌细胞的增殖、迁移和侵袭。 Abstract:ObjectiveTo investigate the mechanism of Periostin (POSTN) regulating AKT signaling pathway to activate the epithelial-mesenchymal transition (EMT) of gastric cancer cells. MethodsPericancerous, cancerous, metastases and primary tumor tissues of patients with gastric cancer at different pathological stage were selected for immunofluorescence staining with POSTN and CD44.POSTN gene was overexpressed or knocked out in human gastric cancer cells MGC 803, and AKT inhibitor LY294002 was added for culture.The expression levels of POSTN, CD44 and EMT-related genes in gastric cancer cells were detected using RT-qPCR and Western blotting.The proliferation level of gastric cancer cells was detected using CCK-8.Transwell assays were used to detect the migration and invasion of cells. ResultsThe expression of POSTN in metastasis was higher than that of primary tumor, and the expression of POSTN in primary tumor was higher than that of pericancerous tissue.Compared with normal MGC 803 cells, the expressions of POSTN, CD44, α-sma, snail, slug, Vimentin were increased and E-Cadherin was reduced in gastric cancer cells overexpressing POSTN, while POSTN knockout inhibited the expressions of CD44, α-sma, snail, slug, Vimentin and increased the expressions of E-Cadherin.The overexpression of POSTN promoted the metastasis and invasion of gastric cancer cells, while the reduction of POSTN inhibited the metastasis and invasion of gastric cancer cells.The addition of LY294002 significantly improved the expression changes of α-sma, snail, slug, Vimentin, E-Cadherin, proliferation activity, migration and invasion ability caused by overexpression of POSTN gene in gastric cancer cells(P < 0.01). ConclusionsThe expression of POSTN in the cancerous and metastatic of gastric cancer is up-regulated, and the later the clinical stage, the higher the expression of POSTN in the cancerous; POSTN activates the EMT of gastric cancer cells by regulating the AKT signaling pathway, thereby promoting the proliferation, migration and invasion of gastric cancer cells. -
Key words:
- gastric neoplasms /
- Periostin /
- epithelial-mesenchymal transition /
- migration /
- invasion
-
表 1 引物序列
基因名称 上游序列 下游序列 GAPDH CTG GGC TAC ACT GAG CAC C AAG TGG TCG TTG AGG GCA ATG CD44 CTG CCG CTT TGC AGG TGT A CAT TGT GGG CAA GGT GCT ATT POSTN CTC ATA GTC GTA TCA GGG GTC G ACA CAG TCG TTT TCT GTC CAC α-sma CCT GTG TTG TGG TTT ACA CTG G GGG GGA ATT ATC TTT CCT GGT CC VIM AGT CCA CTG AGT ACC GGA GAC CAT TTC ACG CAT CTG GCG TTC Snail TCG GAA GCC TAA CTA CAG CGA AGA TGA GCA TTG GCA GCG AG Slug CGA ACT GGA CAC ACA TAC AGT G CTG AGG ATC TCT GGT TGT GGT CDH1 ATT TTT CCC TCG ACA CCC GAT TCC CAG GCG TAG ACC AAG A  下载: 导出CSV
下载: 导出CSV
表 2 CCK8检测调控POSTN基因对MGC 803细胞增殖的影响(x±s)
分组 n OD值 对照组 3 1.06±0.21 过表达组 3 1.80±0.24** 敲除组 3 0.68 ±0.18**△△ F — 25.92 P — < 0.01 MS组内 — 0.045 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
表 3 调控POSTN基因对MGC 803细胞转移的影响(x±s)
分组 n 细胞个数 对照组 3 557.33±125.10 过表达组 3 1 004.67±143.14** 敲除组 3 328.67±94.88**△△ F — 24.67 P — < 0.05 MS组内 — 14 448.000 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
表 4 调控POSTN基因对MGC 803细胞侵袭的影响
分组 n 细胞个数 对照组 3 110.00±44.00 过表达组 3 550.00±166.10** 敲除组 3 55.33±11.02**△△ F — 22.33 P — < 0.05 MS组内 — 9 981.778 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
表 5 RT-qPCR检测POSTN对MGC 803细胞EMT相关基因的影响(x±s)
分组 n POSTN CD44 α-SMA VIM Snail Slug CDH1 对照组 3 1 1 1 1 1 1 1 敲除组 3 0.01±0.01** 0.58±0.32** 0.57±0.17** 0.48±0.15** 0.66±0.10** 0.57±0.31** 1.96±0.38** 过表达组 3 7.66±1.95**△△ 3.30±1.00**△△ 2.88±0.57**△△ 2.67±0.47**△△ 1.88±0.24**△△ 2.10±0.32**△△ 0.55±0.08**△△ F — 41.01 17.48 38.62 49.30 53.48 28.20 31.06 P — < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 1.268 0.369 0.117 0.080 0.022 0.066 0.050 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
表 6 CCK8检测LY294002对MGC 803细胞调控POSTN基因后增殖的影响(x±s)
分组 n OD值 对照组 3 1.28±1.28 LY294002 3 0.61±0.15** 过表达组 3 2.34±0.17** 过表达组+LY294002 3 1.51±0.17**△△ F — 60.23 P — < 0.01 MS组内 — 0.028 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
表 7 LY294002对MGC 803细胞的调控POSTN基因后转移的影响(x±s)
分组 n 细胞个数 对照组 3 412.33±63.12 LY294002 3 269.67±36.23** 过表达组 3 831.33±87.30** 过表达组+LY294002 3 460.33±116.07△△ F — 26.06 P — < 0.01 MS组内 — 6 597.580 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
表 8 LY294002对MGC 803细胞的调控POSTN基因后迁移的影响(x±s)
分组 n 细胞个数 对照组 3 107.67±24.01 LY294002 3 40.00±19.97** 过表达组 3 270.67±67.00** 过表达组+LY294002 3 136.67±37.22△△ F — 16.42 P — < 0.01 MS组内 — 1 712.500 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
表 9 RT-qPCR检测LY294002抑制POSTN激活MGC 803细胞的EMT(x±s)
分组 n POSTN CD44 α-SMA VIM Snail Slug CDH1 AKT 对照组 3 1 1 1 1 1 1 1 1 LY294002 3 1.02±0.18** 0.60±0.16** 0.49±0.15** 0.55±0.11** 0.64±0.07** 0.62±0.10** 1.92±0.64** 0.12±0.08** 过表达组 3 6.45±1.81** 3.34±0.89** 2.92±0.35** 2.82±0.17** 2.14±0.46** 2.17±0.19** 0.52±0.11** 1.03±0.20** 过表达组+LY294002 3 6.30±2.46△△ 1.68±0.39△△ 1.38±0.25△△ 1.22±0.18△△ 1.06±0.09△△ 0.95±0.09△△ 1.11±0.18△△ 0.53±0.11△△ F — 12.32 18.13 64.06 161.55 22.50 101.98 8.89 38.90 P — < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 2.336 0.242 0.051 0.018 0.055 0.014 0.115 0.014 q检验: 与对照组比较**P < 0.01;与过表达组比较△△P < 0.01
下载: 导出CSV
-
[1] SMYTH EC, NILSSON M, GRABSCH HI, et al. Gastric cancer[J]. Lancet, 2020, 396(10251): 635. doi: 10.1016/S0140-6736(20)31288-5 [2] BRAY F, FERLAY J, SOERJOMATARAM I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2018, 68(6): 394. doi: 10.3322/caac.21492 [3] LV YJ, WANG W, JI CS, et al. Association between periostin and epithelial‐mesenchymal transition in esophageal squamous cell carcinoma and its clinical significance[J]. Oncol Lett, 2017, 14(1): 376. doi: 10.3892/ol.2017.6124 [4] SCHWANEKAMP JA, LORTS A, VAGNOZZI RJ, et al. Deletion of periostin protects against atherosclerosis in mice by altering inflammation and extracellular matrix remodeling[J]. Arterioscler Thromb Vasc Biol, 2016, 36(1): 60. doi: 10.1161/ATVBAHA.115.306397 [5] SUN B, GUO W, HU S, et al. Gprc5a-knockout mouse lung epithelial cells predicts ceruloplasmin, lipocalin 2 and periostin as potential biomarkers at early stages of lung tumorigenesis[J]. Oncotarget, 2017, 8(8): 13532. doi: 10.18632/oncotarget.14589 [6] DU B, SHIM JS. Targeting epithelial-mesenchymal transition (EMT) to overcome drug resistance in cancer[J]. Molecules, 2016, 21(7): 965. doi: 10.3390/molecules21070965 [7] FATTAHI S, AMJADI-MOHEB F, TABARIPOUR R, et al. PI3K/AKT/mTOR signaling in gastric cancer: Epigenetics and beyond[J]. Life Sci, 2020, 262: 118513. doi: 10.1016/j.lfs.2020.118513 [8] GONZÁLEZ-GONZÁLEZ L, ALONSO J. Periostin: A matricellular protein with multiple functions in cancer development and progression[J]. Front Oncol, 2018, 8: 225. doi: 10.3389/fonc.2018.00225 [9] NUZZO PV, BUZZATTI G, RICCI F, et al. Periostin: a novel prognostic and therapeutic target for genitourinary cancer?[J]. Clin Genitourin Cancer, 2014, 12(5): 301. doi: 10.1016/j.clgc.2014.02.005 [10] SUAREZ-CARMONA M, LESAGE J, CATALDO D, et al. EMT and inflammation: inseparable actors of cancer progression[J]. Mol Oncol, 2017, 11(7): 805. doi: 10.1002/1878-0261.12095 [11] CHO ES, KANG HE, KIM NH, et al. Therapeutic implications of cancer epithelial-mesenchymal transition (EMT)[J]. Arch Pharm Res, 2019, 42(1): 14. doi: 10.1007/s12272-018-01108-7 [12] WANG L, OUYANG F, LIU X, et al. Overexpressed CISD2 has prognostic value in human gastric cancer and promotes gastric cancer cell proliferation and tumorigenesis via AKT signaling pathway[J]. Oncotarget, 2016, 7(4): 3791. doi: 10.18632/oncotarget.6302 -

 点击查看大图
点击查看大图

图(5)表(9)
计量
- 文章访问数: 2380
- HTML全文浏览量: 1164
- PDF下载量: 23
- 被引次数: 0
