



-
胃癌是最常见的恶性疾病之一,是全世界癌症相关死亡的第三大原因[1-2]。晚期胃癌病人的预后较差,5年总生存率低至约25%[3]。因此,胃癌的早期诊断和有效治疗对人体健康具有重要意义。目前,肿瘤已被广泛认为起源于癌症干细胞。肿瘤干细胞是肿瘤体内的一小部分细胞,具有很高的球形成能力,自我更新、迁移、侵袭以及对化疗和放疗的抵抗力[4]。它们产生不同表型的非致瘤性肿瘤细胞,甚至在治疗结束后导致肿瘤复发。因此,癌症干细胞有助于不同类型肿瘤的发生、发展和复发[5-6]。据报道[7-9],肿瘤干细胞已在多种癌症中成功鉴定,包括黑素瘤、乳腺癌、脑癌、结肠癌和胃癌。新的证据表明[10],肿瘤干细胞是胃癌复发的主要原因,肿瘤干细胞的存在在胃癌进展和治疗抵抗中起关键作用。这些发现表明肿瘤干细胞是胃癌治疗的重要治疗靶点。近年来,星形胶质细胞上调基因-1 (AEG-1)已成为肿瘤进展的几个方面的潜在关键介质[11]。然而,AEG-1在人胃癌干细胞致瘤性中的作用仍不清楚。本文就AEG-1在调节胃癌干细胞中过程中的生物学功能作一探讨。
-
免疫缺陷小鼠(NOD/SCID mice)购于浙江大学动物中心;人胃癌细胞系(HGC-27及MKN-45)由浙江大学生命科学院章晓波教授实验室提供。HGC-27及MKN-45胃癌干细胞使用DMEM/F12培养基(含20 ng/mL表皮生长因子,10 ng/mL碱性成纤维细胞生长因子,5 μg/mL胰岛素和2%的B-27),HGC-27和MKN-45癌非干细胞使用含有10%胎牛血清(FBS)和抗生素的RPMI-1640培养基。以上细胞系均在37 ℃、湿润的5%CO2培养箱中培养。
-
RIPM-1640培养液购自Hyclone公司,DMEM培养液、胎牛血清FBS、胰蛋白酶EDTA-Trpsin购自Gibco公司,转染试剂Lipofectamine 2000购自Invitrogen公司,BCA定量试剂盒购自Byotime公司,CellTiter 96® AQueous One Solution Cell Proliferation Assay (MTS)购自Promega公司。
-
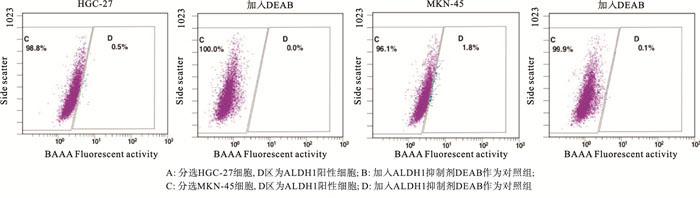
使用ALDEFLUOR试剂盒(Cyagen Biosciences Inc.)进行胃干细胞的分选,以醛脱氢酶1(ALDH1)作为干细胞标志物。将HGC-27或MKN-45细胞悬浮于ALDEFLUOR测定缓冲液中,ALDH1荧光底物BODIPY氨基乙酸酯(BAAA,1 mmol/L)37 ℃孵育40 min。将50 mmol/L二乙基氨基苯甲醛(DEAB)(特异性ALDH1抑制剂)处理等分试样的细胞作为阴性对照。然后将细胞以800 r/min离心5 min。除去上清液后,将细胞沉淀重悬于0.5 mL ALDEFLUOR测定缓冲液中,4 ℃下储存,流式细胞仪荧光激活细胞分选(FACS)。
-
使用RNA Isolation试剂盒从胃癌细胞中分离总RNA,并使用PrimeScript RT Reagent Kit试剂盒进行逆转录。实时PCR检测系统包含cDNA、引物和TaqMan Universal PCR Master Mix。以GAPDH为内参,2-△△CT方法用于测定基因的表达。
-
在6孔板中铺板MKN-45和HGC-27细胞,每孔约60%,使用Lipfectamine2000并用50 nmol/L AEG-1特异性siRNA(AEG-1-siRNA:5′-CAG AAG AAG AAG AAC CGG A-3′)转染,AEG-1-siRNA-scrambled(5′-UUC UCC GAA CGU GUC ACG UTT-3′)作为对照。转染后48 h,收取细胞供后续使用。
-
从细胞中提取蛋白质,加入150 μL裂解液充分裂解,收集上清液,采用BCA法定量并调节蛋白浓度,加入缓冲液煮样10 min,SDS-PAGE分离后转移PVDF膜,然后在室温下用含有5%脱脂乳封闭2 h。随后,将膜与相应的抗体在4 ℃下孵育过夜。TBST溶液洗涤3次后,二抗室温下进一步孵育2 h。在用TBST溶液洗涤后,进行显色。
-
为探索AEG-1 siRNA转染的细胞是否可以抑制胃癌干细胞的生长,我们用AEG-1 siRNA转染HGC-27、MKN-45细胞和对照转染的细胞行MTS测定细胞活力和增殖。将细胞以100 μL培养基以适当密度接种于96孔板中,每个孔中加入20 μL MTS试剂,并在含有5%CO2的培养箱中于37 ℃温育1 h。用酶标仪在450 nm波长下检测每孔的吸光度值,通过绘制剂量效应曲线来进行细胞增殖率分析。每个样品重复3次。
-
为了解AEG-1在胃癌干细胞增殖中的作用机制,检测AEG-1沉默的胃癌干细胞的细胞周期。通过离心收集细胞样品,在预冷的70%乙醇中存放于-20 ℃固定4 h。在800 r/min离心5 min后弃掉乙醇并用PBS清洗细胞3次,用500 μL的PBS重悬细胞,并加入RNase A,37 ℃孵育30 min。用碘化丙锭(PI,50 mg/mL)标记细胞。采用流式细胞仪在488 nm的波长下进行流式细胞术分析。
-
AEG-1基因敲低引起细胞周期阻滞后是否会诱导细胞凋亡发生,我们对AEG-1基因敲除胃癌细胞的细胞凋亡水平进行了检测。转染siRNA后,将细胞以1×104/孔的密度接种于96孔板中。转染48 h后,每孔中加入50 μL Caspase-3/7试剂并在避光的室温下孵育30 min,化学发光检测仪检测其化学发光值。
-
待检测细胞并用预冷的PBS冲洗3次,然后用PBS将10×Annexin V binding buffer母液稀释为1×的工作浓度,向每组待测样品中加入100 μL 1×Annexin V binding buffer重悬。随后,向细胞中加入5 μL Alexa Fluor Annexin V和1 μL的PI。室温避光孵育15 min后,向样品中加入400 μL 1×Annexin V binding buffer,混匀后吸入流式管,用流式细胞仪进行检测。
-
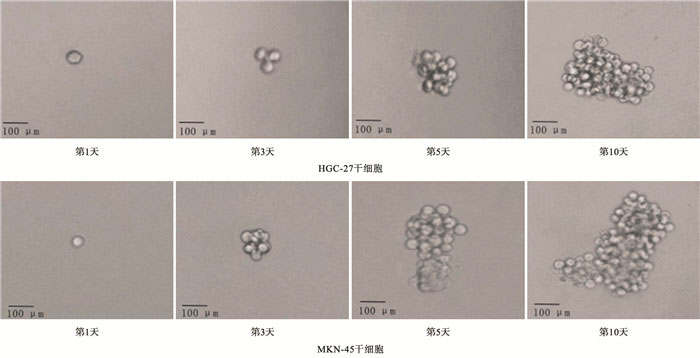
为了验证筛选出的干细胞是否能够在无血清条件下自我更新,将这些分选出的ALDH1阳性细胞进行成球能力测定。将MKN-45、HGC-27两株ALDH1阳性的细胞分别接种一个单个细胞在含有DMEFT12培养基的超黏附96孔板的每个孔中,使用含有2%B-27、20 ng/mL表皮细胞生长因子、10 ng/mL成纤维细胞生长因子及5 μg/mL胰岛素的培养基,在第1、3、5和10天通过光学显微镜检查成球情况。
-
为了评估体内ALDH1阳性细胞的肿瘤形成能力,将2株细胞分别计数500个ALDH1阳性的细胞皮下注射到免疫缺陷小鼠中,6周后处死裸鼠可见瘤块形成。为了探索AEG-1对体内肿瘤生长的影响,通过离心收集胃癌干细胞,计数4 000个细胞,用2 mL 0.9%氯化钠溶液重悬后注射到裸鼠背部,每只0.2 mL,观察肿瘤生长。3周后,当肿瘤体积约为20 mm3时,每3天用80 mg/kg AEG-1-siRNA或AEG-1-siRNA-scrambled注射小鼠尾静脉,每周测量肿瘤体积,肿瘤体积=(长径×短径×短径)/2,其中宽度和长度分别是最短和最长直径。在距第1次注射第55天,通过颈脱位处死小鼠,解剖肿瘤并称重。所有动物实验均按照IACUC批准的方案进行。
-
采用t检验。
-
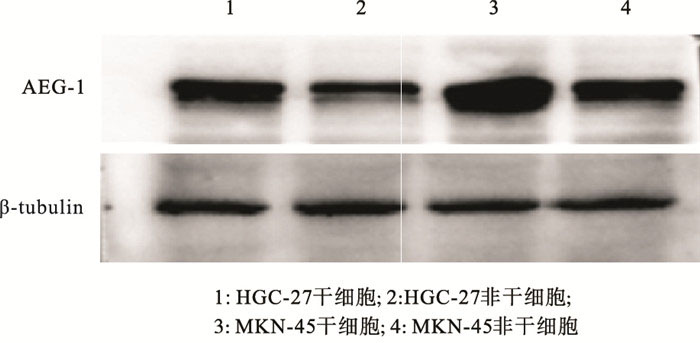
ALDH1阳性细胞是肿瘤干细胞,而ALDH1阴性细胞是非干细胞(见图 1)。ALDH1阳性细胞均能形成大的细胞球(见图 2)。注射ALDH1阳性细胞的裸鼠中都能长成肿瘤(见图 3)。定量实时PCR的结果显示,与癌症非干细胞相比,AEG-1表达在胃癌干细胞MKN-45中显著上调(P < 0.01)(见表 1)。Western blotting基本上也显示了类似的结果(见图 4)。

图 1 胃癌干细胞的分选
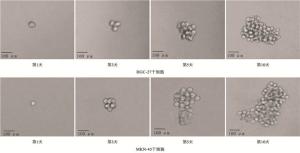
图 2 ALDH1阳性细胞成球能力测定

图 3 ALDH1阳性细胞肿瘤形成能力
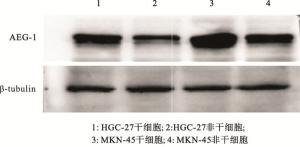
图 4 Western blotting检测AEG-1在胃癌干细胞及非干细胞中蛋白表达
分组 n HGC-27 MKN-45 干细胞 3 1.083±0.055 1.100±0.089 非干细胞 3 0.620±0.046 0.753±0.055 t — 1.45* 5.74 P — >0.05 < 0.01 *示t′值 表 1 定量PCR分析AEG-1的mRNA水平(x±s)
-
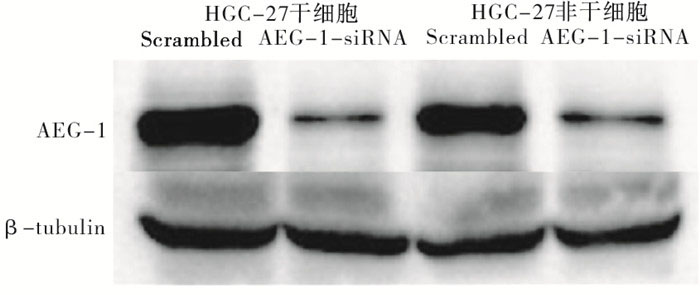
当将AEG-1特异性siRNA转染到胃癌干细胞中时,mRNA表达非常低(P < 0.01)(见表 2)。进一步通过Western blotting分析检测蛋白质表达,与对照相比,低mRNA表达与低蛋白质表达相关(见图 5)。AEG-1 siRNA转染的胃癌干细胞增殖明显减少(P < 0.01),而AEG-1敲低后对胃癌非干细胞的生长没有显著影响(P>0.05)(见表 3)。AEG-1-siRNA处理的细胞在G1期中的百分比远高于对照组(P < 0.01)(见表 4)。
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.197±0.019 0.200±0.090 0.155±0.043 0.189±0.065 AEG-1-siRNA-scrambled 3 1.045± 0.059 0.616±0.045 1.105±0.085 0.765±0.059 t — 23.68 7.16 17.27 11.37 P — < 0.01 < 0.01 < 0.01 < 0.01 表 2 AEG-1-siRNA转染HGC-27或MKN-45干细胞和非干细胞中的AEG-1 mRNA水平(x±s)
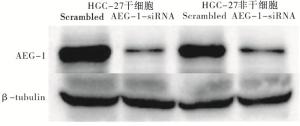
图 5 Western blotting检测敲低AEG-1后在胃癌干细胞及非干细胞中蛋白表达
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.669±0.054 1.108±0.056 0.740±0.075 1.097±0.075 AEG-1-siRNA-scrambled 3 1.288±0.115 1.097±0.049 1.382±0.019 1.118±0.083 t — 8.44 0.26 13.52 0.33 P — < 0.01 >0.05 < 0.01 >0.05 表 3 AEG-1敲低对HGC-27或MKN-45干细胞及非干细胞增殖的影响(x±s)
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.510±0.023 0.381±0.020 0.412±0.032 0.408±0.026 AEG-1-siRNA-scrambled 3 0.379±0.026 0.403±0.0.007 0.274±0.021 0.428±0.026 t — 6.52 1.79 6.24 0.94 P — < 0.01 >0.05 < 0.01 >0.05 表 4 AEG-1敲低对HGC-27或MKN-45干细胞及非干细胞细胞周期的影响(x±s)
-
Caspase-3/7活性检测显示,与对照相比,胃癌干细胞中AEG-1基因敲除后的细胞凋亡比例显著上升(P < 0.01)(见表 5)。Annexin V实验发现,AEG-1敲低可诱导胃干细胞凋亡(P < 0.01)(见图 6、表 6)。
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 9 385.7±520.5 6 126.0±788.0 8 513.3±575.0 6 019.7±934.1 AEG-1-siRNA-scrambled 3 5 775.0±549.2 5 964.7±788.3 5 619.0±1 252.5 5 664.3±414.2 t — 8.27 0.25 3.64 0.60 P — < 0.01 >0.05 < 0.05 >0.05 表 5 AEG-1基因敲低对胃癌细胞Caspase-3/7酶活的影响(x±s)

图 6 Annexin V检测敲低AEG-1后胃癌干细胞凋亡的结果
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.163±0.012 0.043±0.006 0.173±0.006 0.067±0.015 AEG-1-siRNA-scrambled 3 0.047±0.006 0.033±0.006 0.037±0.006 0.047±0.006 t — 14.98 2.04 27.76 2.14 P — < 0.01 >0.05 < 0.01 >0.05 表 6 Annexin V检测AEG-1基因敲低胃癌细胞的细胞凋亡情况(x±s)
-
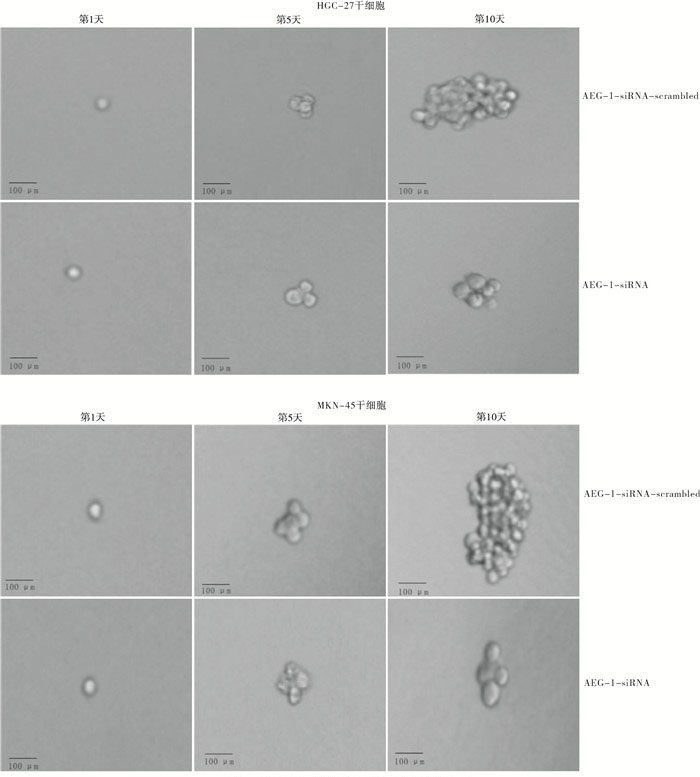
当AEG-1表达被敲低时,显著抑制了胃癌干细胞的成球能力,其长成的细胞球明显小于对照组(见图 7)。
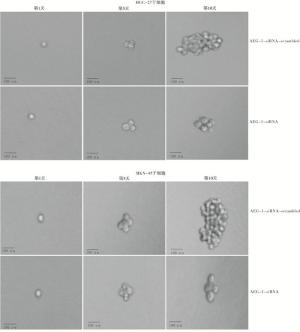
图 7 敲低AEG-1对胃癌干细胞成球能力的影响
-
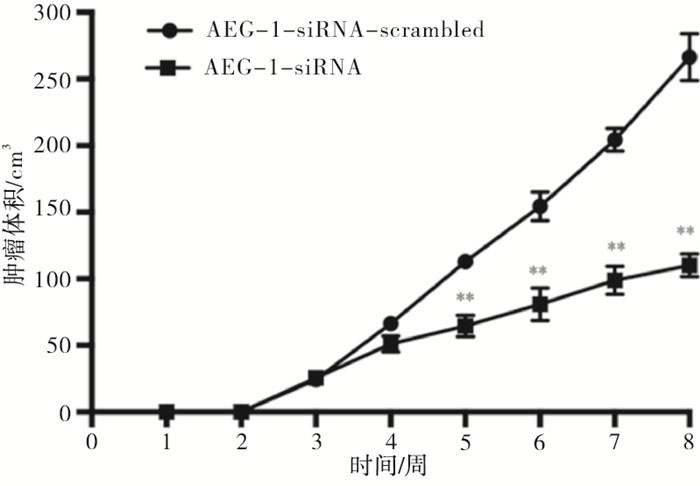
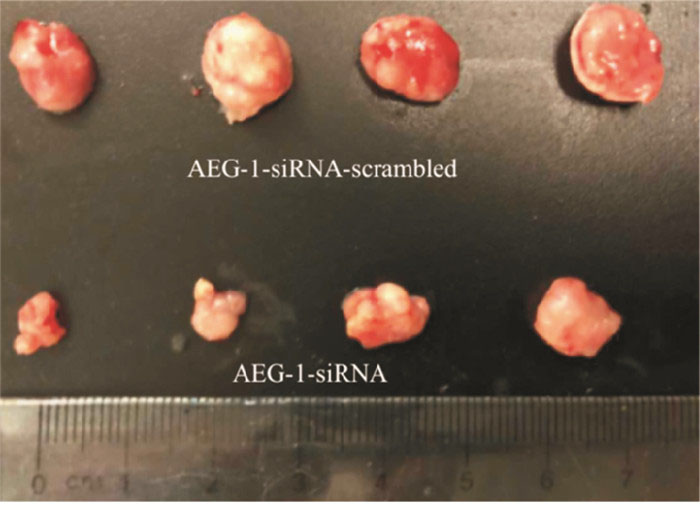
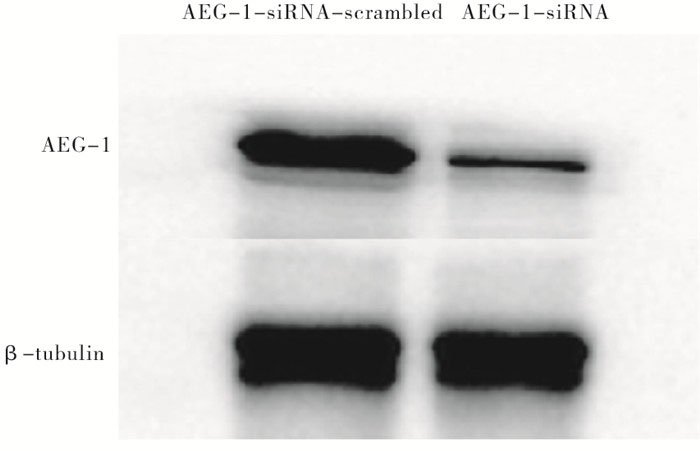
2组处理的瘤块大小及瘤块质量存在显著差异(见图 8),注射AEG-1-siRNA处理的小鼠肿瘤瘤块体积和质量(0.167±0.042)g明显小于对照组(0.480±0.030)g(t=10.50,P < 0.01)(见图 9)。采用Western blotting检测瘤块内AEG-1的表达情况,与对照组相比,AEG-1敲低显著降低了小鼠实体瘤中AEG-1的表达水平(见图 10)。
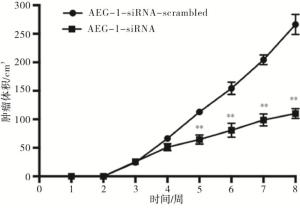
图 8 敲低AEG-1对小鼠瘤块体积的影响
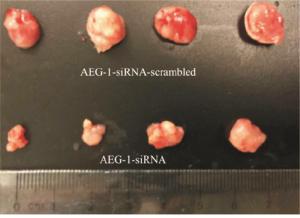
图 9 敲低AEG-1对小鼠瘤块的影响

图 10 Western blotting检测裸鼠的瘤块内AEG-1蛋白水平(β-tubulin作为试验的对照)
-
肿瘤干细胞属于肿瘤细胞群中一类功能不同的细胞亚型,具有自我更新、多向分化、高致瘤性和对放疗和化疗抗性的生物学特征。肿瘤干细胞可以在补充有生长因子的无血清悬浮培养基中培养形成球状细胞,并保持自我更新特性,而非干细胞则不能存活。该技术被认为是获得癌干细胞的一种便捷方式[12]。HAN等[13-14]使用这种方法从胃癌组织和胃癌细胞系中鉴定了胃癌干细胞,并且当注射到严重联合免疫缺陷小鼠的胃和皮肤中时致肿瘤形成,表明具有有效的体内肿瘤起始能力。这些肿瘤干细胞表现干细胞样特性,包括持续自我更新、多向分化和高致瘤性,导致即使根治性手术和术后辅助治疗后,胃癌疾病仍预后不良[15]。近年来,AEG-1已成为各种器官和组织中致癌过程的关键因素[16]。AEG-1表达在涉及人体几乎所有部分的多种癌症中升高[16-17]。AEG-1过表达与癌症临床预后不良相关[18]。据报道[19-20],AEG-1可通过激活核因子κ-B(NF-κB)、磷脂酰肌醇3-激酶(PI3K)、c-Myc和Wnt/β-catenin来促进肿瘤增殖、侵袭、化疗耐药、血管生成和转移。AEG-1的上调可以诱导NF-κB的转录活性和细胞质/细胞核转位,NF-κB的抑制显着逆转了Hela细胞和人胶质瘤细胞中AEG-1诱导的琼脂克隆效率和基质胶侵袭[21]。AEG-1的异常表达通过NF-κB和PI3K-AKT途径影响非小细胞肺癌细胞系的迁移和侵袭[22]。重组AEG-1可激活Wnt信号通路,其对舌细胞侵袭性和上皮-间质转化的鳞状细胞癌的刺激作用被抗Wnt5a中和抗体或通过抑制Rac1或ROCK逆转[23]。AEG-1表达不仅在调节致癌转化中起主导作用,而且还参与血管生成过程。AEG-1的过表达增强血管生成因子的产生,包括血管生成素-1、基质金属蛋白酶-2、Tie2和缺氧诱导因子1-α[24]。越来越多的证据[25]表明AEG-1在胃癌中发挥重要作用,已发现AEG-1在胃癌组织中过表达,AEG-1的下调可以抑制胃癌细胞的生长和肿瘤的进展。然而,到目前为止,AEG-1对胃癌干细胞的影响尚未探索。研究[26]表明,肿瘤由异质细胞群组成,这些细胞群的相对分化状态不同,在肿瘤的核心区域是癌干细胞,在结构和功能上都与肿瘤块中的其他细胞不同。癌症干细胞负责肿瘤的发生、转移和治疗抵抗。因此,AEG-1在胃癌干细胞中的作用值得研究。本文结果显示,AEG-1沉默抑制细胞增殖,诱导胃癌干细胞G1期细胞周期停滞和细胞凋亡,从而抑制肿瘤发生,提示AEG-1在胃癌干细胞的肿瘤发生中起着积极的作用,AEG-1可能是治疗胃癌的新型治疗靶点。在未来的研究中,AEG-1沉默对胃癌细胞转移的影响值得探讨。
恶性肿瘤的增殖是获得无限生长潜力和抗凋亡的结果。已有报道[27]AEG-1通过抑制血清饥饿诱导的细胞凋亡来增强血清非依赖性细胞生长。AEG-1的敲低导致细胞周期的G0/G1期细胞停滞,从而抑制集落形成能力,并促进神经母细胞瘤细胞的凋亡,而AEG-1过表达与Ki67的高水平显著相关,Ki67是一种常见的增殖标志物[28]。在卵巢癌中,AEG-1的低表达可以抑制癌细胞的增殖,诱导细胞凋亡并降低癌细胞的侵袭能力[29]。还有报道[30]AEG-1通过介导miR-154表达抑制胃癌细胞的增殖。本研究也显示AEG-1沉默可导致癌症干细胞周期停滞在G1期,表明AEG-1对胃癌干细胞的增殖具有积极作用。
本研究也证实AEG-1基因敲低后会诱导肿瘤干细胞发生细胞凋亡,并抑制肿瘤干细胞的体内成瘤能力,表明AEG-1在维持胃癌干细胞干性方面起着关键作用。据报道[31],AEG-1表达的抑制通过PI3K/Akt信号通路促进前列腺癌细胞中的FOXO3a活性和乳腺癌细胞中FOXO1的活性,从而诱导癌细胞凋亡。在非小细胞肺癌中,AEG-1显著增加PI3Kp110的水平和Akt的磷酸化,导致PI3K/Akt通路的激活,并通过抑制Caspase-3和增强抗凋亡蛋白Bcl-2的水平来抑制细胞凋亡[32]。在胃癌中,AEG-1的抑制降低了β-连环蛋白的水平,并下调了Wnt/β-catenin通路中两个关键的下游效应因子淋巴增强因子1和细胞周期蛋白D1蛋白, 从而抑制肿瘤细胞生长并增强细胞凋亡[29]。表明在不同癌症中AEG-1通过PI3K/Akt信号通路或Wnt/β-连环蛋白途径参与癌细胞的凋亡。另据报道[33],血管生成是实体瘤及其转移的维持和发展中的基本事件,血管生成的过程受促血管生成因子的相互作用调节,AEG-1在调节血管生成中起主导作用[33-34],由此推测,AEG-1通过调节PI3K/Akt信号转导途径、Wnt/β-连环蛋白途径和/或肿瘤细胞的血管生成而在肿瘤发生中起作用。AEG-1介导的胃癌干细胞癌进展的机制值得进一步研究。
星形胶质细胞上调基因-1对胃癌干细胞增殖、凋亡及干性的影响
Effect of astrocyte elevated gene-1 on the proliferation, apoptosis and stemness of gastric cancer stem cells
-
摘要:
目的探讨星形胶质细胞上调基因-1(AEG-1)在胃癌干细胞中的表达及其作用。 方法采用实时荧光定量技术(qRT-PCR)及Western blotting检测胃癌干细胞(HGC-27、MKN-45)及相应非干细胞mRNA及蛋白表达水平,采用瞬时转染法对AEG-1进行处理,使用MTS、细胞周期检测细胞增殖情况及作用机制,Annexin Ⅴ及Caspase-3/7活性检测细胞凋亡情况,干细胞成球实验评估AEG-1沉默对胃干细胞干性的影响,裸鼠体内成瘤实验评估AEG-1对裸鼠体内胃干细胞肿瘤发生的影响。Ⅴ 结果胃癌干细胞MKN-45中AEG-1表达水平高于非干细胞组(P < 0.01);AEG-1-siRNA转染的癌干细胞导致癌干细胞周期停滞在G1期,明显抑制了细胞增殖(P < 0.01);Annexin Ⅴ及Caspase 3/7活性检测AEG-1敲低可诱导胃干细胞凋亡;AEG-1表达被敲低时显著抑制了胃癌干细胞的成球能力;AEG-1缺失抑制了肿瘤干细胞的体内成瘤能力。 结论AEG-1沉默抑制细胞增殖,诱导胃癌干细胞G1期细胞周期停滞和细胞凋亡,抑制肿瘤发生,在胃癌干细胞的肿瘤发生中起着积极作用。 -
关键词:
- 胃肿瘤 /
- 星形胶质细胞上调基因-1 /
- 肿瘤干细胞
Abstract:ObjectiveTo investigate the expression level and effects of astrocyte elevated gene-1(AEG-1) in gastric cancer stem cells. MethodsThe expression levels of AEG-1 mRNA and protein in gastric stem cell lines(HGC-27 and MKN-45) were detected using RT-PCR and Western blotting, respectively.The AEG-1 was processed using transient transfection, and the cells proliferation and its mechanism were investigated using the MTS assay and cell-cycle analysis.The apoptosis was examined using the activities of Caspase-3/7 and annexin V, the effect of silencing AEG-1 on the stemness of gastric stem cells was evaluated using the tumorsphere formation assay, and the tumor formation experiment was conducted to evaluate the effects of AEG-1 on the tumorigenesis of gastric stem cells in nude mice. ResultsThe expression level of AEG-1 in gastric stem cells was higher than that in non-stem cells(P < 0.01).Cancer stem cells transfected with AEG1-siRNA resulted in the cancer stem cell cycle arrest in G1 phase, and could significantly inhibit the cell proliferation(P < 0.01).The results of Annexin V and Caspase-3/7 activities detection showed that the knockdown of AEG-1 could induce the apoptosis of gastric stem cells.The low level of AEG-1 could significantly inhibit the ability of gastric cancer stem cells to form into balls.The loss of AEG-1 inhibited the tumorigenesis of tumor stem cells in vivo. ConclusionsThe silencing expression of AEG-1 can suppress the cell proliferation, induce the cell cycle arrest in G1 phase and apoptosis of gastric cancer stem cells, inhibit the tumorigenesis, which plays a positive role in tumorigenesis of gastric cancer stem cells. -
Key words:
- gastric neoplasms /
- astrocyte elevated gene-1 /
- cancer stem cell
-
表 1 定量PCR分析AEG-1的mRNA水平(x±s)
分组 n HGC-27 MKN-45 干细胞 3 1.083±0.055 1.100±0.089 非干细胞 3 0.620±0.046 0.753±0.055 t — 1.45* 5.74 P — >0.05 < 0.01 *示t′值  下载: 导出CSV
下载: 导出CSV
表 2 AEG-1-siRNA转染HGC-27或MKN-45干细胞和非干细胞中的AEG-1 mRNA水平(x±s)
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.197±0.019 0.200±0.090 0.155±0.043 0.189±0.065 AEG-1-siRNA-scrambled 3 1.045± 0.059 0.616±0.045 1.105±0.085 0.765±0.059 t — 23.68 7.16 17.27 11.37 P — < 0.01 < 0.01 < 0.01 < 0.01
下载: 导出CSV
表 3 AEG-1敲低对HGC-27或MKN-45干细胞及非干细胞增殖的影响(x±s)
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.669±0.054 1.108±0.056 0.740±0.075 1.097±0.075 AEG-1-siRNA-scrambled 3 1.288±0.115 1.097±0.049 1.382±0.019 1.118±0.083 t — 8.44 0.26 13.52 0.33 P — < 0.01 >0.05 < 0.01 >0.05
下载: 导出CSV
表 4 AEG-1敲低对HGC-27或MKN-45干细胞及非干细胞细胞周期的影响(x±s)
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.510±0.023 0.381±0.020 0.412±0.032 0.408±0.026 AEG-1-siRNA-scrambled 3 0.379±0.026 0.403±0.0.007 0.274±0.021 0.428±0.026 t — 6.52 1.79 6.24 0.94 P — < 0.01 >0.05 < 0.01 >0.05
下载: 导出CSV
表 5 AEG-1基因敲低对胃癌细胞Caspase-3/7酶活的影响(x±s)
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 9 385.7±520.5 6 126.0±788.0 8 513.3±575.0 6 019.7±934.1 AEG-1-siRNA-scrambled 3 5 775.0±549.2 5 964.7±788.3 5 619.0±1 252.5 5 664.3±414.2 t — 8.27 0.25 3.64 0.60 P — < 0.01 >0.05 < 0.05 >0.05
下载: 导出CSV
表 6 Annexin V检测AEG-1基因敲低胃癌细胞的细胞凋亡情况(x±s)
分组 n HGC-27 MKN-45 干细胞 非干细胞 干细胞 非干细胞 AEG-1-siRNA 3 0.163±0.012 0.043±0.006 0.173±0.006 0.067±0.015 AEG-1-siRNA-scrambled 3 0.047±0.006 0.033±0.006 0.037±0.006 0.047±0.006 t — 14.98 2.04 27.76 2.14 P — < 0.01 >0.05 < 0.01 >0.05
下载: 导出CSV
-
[1] CHEN YL, ZHANG SY, WANG QZ, et al.Tumor-recruited M2 macrophages promote gastric and breast cancer metastasis via M2 macrophage-secreted CHI3L1 protein[J].J Hemato Oncol, 2017, 10:36. doi: 10.1186/s13045-017-0408-0 [2] PARK JY, VON KARSA L, HERRERO R.Prevention strategies for gastric cancer:a global perspective[J].Clin Endosc, 2014, 47(6):478. doi: 10.5946/ce.2014.47.6.478 [3] HUANG Z, ZHU D, WU L, et al.Six serum-based miRNAs as potential diagnostic biomarkers for gastric cancer[J].Cancer Epidemiol Biomarkers Prev, 2017, 26(2):188. doi: 10.1158/1055-9965.EPI-16-0607 [4] SHAO QW, XU JING, GUAN XIN, et al.In vitro and in vivo effects of miRNA-19b/20a/92a on gastric cancer stem cells and the related mechanism[J].Int J Med Sci, 2018, 15(1):86. doi: 10.7150/ijms.21164 [5] HAN ME, JEON TY, HWANG SH, et al.Cancer spheres from gastric cancer patients provide an ideal model system for cancer stem cell research[J].Cell Mol Life Sci, 2011, 68(21):3589. doi: 10.1007/s00018-011-0672-z [6] COLAK S, MEDEMA JP.Cancer stem cells-important players in tumor therapy resistance[J].FEBS J, 2014, 281(21):4779. doi: 10.1111/febs.13023 [7] BOCKHORN J, YEE K, CHANG YF, et al.MicroRNA-30c targets cytoskeleton genes involved in breast cancer cell invasion[J].Breast Cancer Res Treat, 2013, 137(2):373. doi: 10.1007/s10549-012-2346-4 [8] LIN CW, CHANG YL, CHANG YC, et al.MicroRNA-135b promotes lung cancer metastasis by regulating multiple targets in the Hippo pathway and LZTS1[J].Nat Commun, 2013, 4:1877. doi: 10.1038/ncomms2876 [9] YANG F, WEI J, ZHANG S, et al.Shrimp miR-S8 suppresses the stemness of human melanoma stem-like cells by targeting the transcription factor YB-1[J].Cancer Res, 2017, 77(20):5543. doi: 10.1158/0008-5472.CAN-17-1375 [10] ZHANG XW, HUA RX, WANG XF, et al.Identification of stem-like cells and clinical significance of candidate stem cell markers in gastric cancer[J].Oncotarget, 2016, 7(9):9815. [11] LAWRENCE N, CHIN KL.The metastasis problem gets stickier[J].Cancer Cell, 2009, 15(1):1. doi: 10.1016/j.ccr.2008.12.007 [12] GRIMSHAW MJ, COOPER L, PAPAZISIS K, et al.Mammosphere culture of metastatic breast cancer cells enriches for tumorigenic breast cancer cells[J].Breast Cancer Res, 2008, 10(3):R52. [13] HAN ME, JEON TY, HWANG SH, et al.Cancer spheres from gastric cancer patients provide an ideal model system for cancer stem cell research[J].Cell Mol Life Sci, 2011, 68(21):3589. doi: 10.1007/s00018-011-0672-z [14] SONG Z, YUE W, WEI B, et al.Sonic hedgehog pathway is essential for maintenance of cancer stem-like cells in human gastric cancer[J].PLoS One, 2011, 6(3):e17687. doi: 10.1371/journal.pone.0017687 [15] JEMAL A, BRAY F, CENTER MM, et al.Global cancer statistics[J].CA Cancer J Clin, 2011, 61(2):69. doi: 10.3322/caac.v61:2 [16] HUANG Y, LI LP.Progress of cancer research on astrocyte elevated gene-1/Metadherin (Review)[J].Oncol Lett, 2014, 8(2):493. [17] SHI X, WANG X.The role of MTDH/AEG-1 in the progression of cancer[J].Int J Clin Exp Med, 2015, 8(4):4795. [18] CHEN X, DONG K, LONG M, et al.Serum anti-AEG-1 autoantibody is a potential novel biomarker for malignant tumors[J].Oncol Lett, 2012, 4(2):319. doi: 10.3892/ol.2012.734 [19] LIU H, SONG X, LIU C, et al.Knockdown of astrocyte elevated gene-1 inhibits proliferation and enhancing chemo-sensitivity to cisplatin or doxorubicin in neuroblastoma cells[J].J Exp Clin Cancer Res, 2009, 28:19. doi: 10.1186/1756-9966-28-19 [20] EMDAD L, LEE SG, SU ZZ, et al.Astrocyte elevated gene-1(AEG-1) functions as an oncogene and regulates angiogenesis[J].Proc Natl Acad Sci USA, 2009, 106(50):21300. doi: 10.1073/pnas.0910936106 [21] SARKAR D, PARK ES, EMDAD L, et al.Molecular basis of nuclear factor-kappa B activation by astrocyte elevated gene-1[J].Cancer Res, 2008, 68(5):1478. doi: 10.1158/0008-5472.CAN-07-6164 [22] SONG L, LI W, ZHANG H, et al.Over-expression of AEG-1 significantly associates with tumour aggressiveness and poor prognosis in human non-small cell lung cancer[J].J Pathol, 2009, 219(3):317. [23] PAN Y, GUO X, YANG Z, et al.AEG-1 activates Wnt/PCP signaling to promote metastasis in tongue squamous cell carcinoma[J].Oncotarget, 2016, 7(2):2093. [24] WEI Y, HU G, KANG Y.Metadherin as a link between metastasis and chemoresistance[J].Cell Cycle, 2009, 8(14):2132. [25] HUANG W, YANG L, LIANG S, et al.AEG-1 is a target of perifosine and is over-expressed in gastric dysplasia and cancers[J].Dig Dis Sci, 2013, 58(10):2873. doi: 10.1007/s10620-013-2735-5 [26] HATFIELD S, RUOHOLA-BAKER H.microRNA and stem cell function[J].Cell Tissue Res, 2008, 331(1):57. [27] LEE SG, SU ZZ, EMDAD L, et al.Astrocyte elevated gene-1 activates cell survival pathways through PI3K-Akt signaling[J].Oncogene, 2008, 27(8):1114. doi: 10.1038/sj.onc.1210713 [28] SU P, ZHANG Q, YANG Q.Immunohistochemical analysis of Metadherin in proliferative and cancerous breast tissue[J].Diagn Pathol, 2010, 5:38. doi: 10.1186/1746-1596-5-38 [29] WANG J, CHEN X, TONG M.Knockdown of astrocyte elevated gene-1 inhibited cell growth and induced apoptosis and suppressed invasion in ovarian cancer cells[J].Gene, 2017, 616:8. doi: 10.1016/j.gene.2017.03.024 [30] QIAO W, CAO N, YANG L.microRNA 154 inhibits the growth and metastasis of gastric cancer cells by directly targeting MTDH[J].Oncol Lett, 2017, 14(3):3268. doi: 10.3892/ol.2017.6558 [31] LI J, YANG L, SONG L, et al.Astrocyte elevated gene-1 is a proliferation promoter in breast cancer via suppressing transcriptional factor FOXO1[J].Oncogene, 2009, 28(36):3188. doi: 10.1038/onc.2009.171 [32] KE ZF, MAO X, ZENG C, et al.AEG-1 expression characteristics in human non-small cell lung cancer and its relationship with apoptosis[J].Med Oncol, 2013, 30(1):383. doi: 10.1007/s12032-012-0383-9 [33] CARMELIET P, JAIN RK.Angiogenesis in cancer and other diseases[J].Nature, 2000, 407(6801):249. doi: 10.1038/35025220 [34] SRIVASTAVA J, SIDDIQ A, EMDAD L, et al.Astrocyte elevated gene-1 promotes hepatocarcinogenesis:novel insights from a mouse model[J].Hepatology, 2012, 56(5):1782. doi: 10.1002/hep.25868 -

 点击查看大图
点击查看大图
图(10)表(6)
计量
- 文章访问数: 6169
- HTML全文浏览量: 3475
- PDF下载量: 22
- 被引次数: 0
