



-
非酒精性脂肪肝(NAFLD)与胰岛素抵抗、糖尿病、代谢综合征和心血管疾病发病密切有关[1],严重时可能发展为肝硬化、肝癌。生活方式干预为治疗的首选,目前仍缺乏较为有效的治疗药物。胰高血糖素样肽-1(GLP-1)是一种由小肠L细胞分泌的具有刺激胰岛素分泌功能的激素,能调节食物摄入、胃肠蠕动、体液稳态和脂质代谢,刺激细胞增殖,减少炎症和细胞凋亡[2]。利拉鲁肽(LG)是GLP-1的类似物,2009年首次获得许可用于肥胖2型糖尿病(T2DM)病人的血糖控制。LG具有抑制中枢食欲、延迟胃排空时间和诱导剂量依赖性体质量减轻的特性,以往的研究表明LG可以通过非减肥的方式减轻高脂饮食(HFD)诱导的肝脏脂质积聚[3]。LG成为NAFLD病人考虑的一种有希望的治疗选择,然而,LG对NAFLD影响的机制尚不清楚。
腺苷酸活化蛋白激酶(AMPK)/雷帕霉素靶蛋白(mTOR)是调节自噬的主要通路[4],激活AMPK / mTOR通路调控自噬是一种重要的保护机制[5]。研究[6]表明,LG可以减轻肝脏脂肪变性,减少氧化应激,并激活自噬。LG除了改善T2DM和NAFLD病人的血糖控制外,还能降低体质量、肝内脂肪和内脏脂肪含量[7]。本研究基于AMPK/mTOR通路探讨LG在肝细胞脂肪变性的影响,观察自噬调节蛋白的变化。
-
LG为诺和诺德(中国制药有限公司生产;AMPK抑制剂(Compound C)购于英国Abcam公司;免疫染色固定液、一抗、二抗、RIPA裂解液、Western转膜液、BCA蛋白浓度测定试剂盒、SDS-PAGE凝胶配制试剂盒和SDS-PAGE电泳液均购于碧云天生物技术研究所;PBS缓冲液购于以色列BI生物公司;胎牛血清购于美国GIBCO公司;全自动生化分析仪购于美国贝克曼公司;普通显微镜购于日本Nikon公司;荧光显微镜购于日本OLYMPUS公司;组织包埋机(EG11508)、石蜡切片机(RM2135)购于德国Leica公司;Western blotting用电泳槽(DYCZ-24DN)、转移电泳仪(DYCZ-24DN)及恒温循环器(WD-9412A)、制冰机(SIM-F124)均购于北京六一生物科技有限公司。
-
SPF级雄性C57BL/6小鼠30只,购于北京维通利华有限公司,7~8周龄,体质量(25.2±3.5)g。将小鼠随机分为对照组(n=10)、模型组(n=10)和LG组(n=10),后2组应用高脂饲料(脂肪56.0%、蛋白质7.0%、糖类37.0%)喂养建立肝脂肪变性模型,对照组应用普通饲料(脂肪10.2%、蛋白质23.3%、糖类66.5%)喂养。12周后,LG组应用LG腹腔注射0.6 mg·kg-1·d-1,其他2组应用等量0.9%氯化钠溶液,连续用药4周。
-
应用含0.3 mmol/L棕榈酸(PA)的培养基培养24 h诱导建立HepG2肝细胞脂肪变性模型后,将细胞等量分为模型组(PA)、LG组(PA+LG)和AMPK抑制剂组(PA+LG+Compound C)。LG组加入100 nmol/L LG,AMPK抑制剂组加入100 nmol/L LG和Compound C 20 μmol/L。待细胞长满后进行实验。
-
小鼠禁食、禁水12 h,10%水合氯酸麻醉后,摘取双眼球取血,3 000 r/min离心25 min,于-80 ℃保存。应用全自动生化分析仪检测丙氨酸氨基转移酶(ALT)、天冬氨酸氨基转移酶(AST)、低密度脂蛋白(LDL)、胆固醇(TC)、三酰甘油(TG)。
-
石蜡切片,常规脱蜡。苏木精染色、浸洗、1%盐酸与乙醇混合溶液浸泡、流水冲洗返蓝、蒸馏水过洗、5%伊红染色,最后脱水、透明、封片。显微镜观察肝细胞组织形态和脂肪变性。
-
冰冻切片,蒸馏水充分洗涤,油红O染色10 min。乙醇分化,蒸馏水洗1~2 s,苏木素复染核,蒸馏水洗1~2 s,封片。显微镜观察肝细胞内脂质沉积的情况。
-
提取肝脏总蛋白后,通过BCA法测定蛋白质浓度。按照比例加入5×电泳加样缓冲液并置于沸水浴中10 min。按每孔30 μg蛋白量上样,进行SDS-PAGE凝胶电泳(浓缩胶80 V/40 min,分离胶110 V/60 min),结束电泳后,将分离的蛋白转膜至PVDG膜(300 mA),5%脱脂奶粉封闭,加入一抗4 ℃过夜,洗膜后加入二抗,室温孵育1 h洗膜,应用ECL试剂盒显影,并进行统计分析。
-
将贴壁的HepG2细胞胰酶消化后制成细胞悬液,将无菌细胞爬片放置于六孔板底部后滴加500 μL细胞悬液,置于5%CO2细胞培养箱孵育2 h使HepG2细胞贴于细胞爬片上;每孔加1 200 μL 2%FBS培养基,继续孵育8 h。爬片后,双氧水孵育30 min,含有Tween-20的3%牛血清白蛋白/ PBS缓冲液封闭2 h,一抗过夜孵育,再用PBS缓冲液清洗2次。二抗孵育30 min,用PBS缓冲液清洗2次,再用1 μg/mL DAPI复染。所有的操作步骤都在室温下进行。在100×的油镜下使用荧光显微镜观察并采集图像。
-
提取细胞总蛋白,按照1.3.4的实验方法检测HepG2细胞p-AMPK、p-mTOR、LC3B、Beclin1的蛋白表达。
-
采用方差分析和q检验。
-
结果显示,3组之间总体均数差异有统计学意义,其中模型组与对照组比较,小鼠血清ALT、AST、LDL、TC、TG升高; LG组与模型组相比,血清ALT、AST、LDL、TC、TG下降,差异具有统计学意义(P < 0.05~P < 0.01)(见表 1)。
分组 n ALT/(U/L) AST/(U/L) LDL/(mmo/L) TC/(mmol/L) TG/(mmol/L) 对照组 10 11.33±1.75 31.04±4.26 0.44±0.09 2.04±0.17 0.34±0.05 模型组 10 107.80±7.33** 63.49±8.90** 1.04±0.15** 5.02±0.55** 1.22±0.18** LG组 10 40.40±3.05**## 40.27±6.02**## 0.57±0.12*## 3.57±0.45**## 0.46±0.11*## F — 1 111.98 62.79 68.54 126.41 144.35 P — < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 22.022 44.523 0.015 0.176 0.016 q检验:与对照组相比*P < 0.05,**P < 0.01;与模型组相比##P < 0.01 表 1 各组小鼠ALT、AST、LDL、TC、TG比较(x±s)
-
结果显示,对照组小鼠肝细胞排列规则、细胞核清晰、细胞质均匀、基本无脂肪变性,模型组肝细胞排列紊乱、细胞膨胀、可见大量脂肪空泡形成,LG组较模型组相比,肝细胞排列较其整齐,脂肪空泡明显减少。说明LG能改善肝细胞脂肪变性(见图 1)。

图 1 小鼠肝组织肝细胞形态变化(HE染色)
-
结果显示,对照组小鼠肝组织细胞中偶见橘黄色脂滴,高脂喂养后模型组小鼠肝组织细胞可见大量橘黄色脂滴,融合成片,而LG干预后脂滴含量明显减少(见图 2)。

图 2 小鼠肝细胞脂肪滴变化(油红O染色)
-
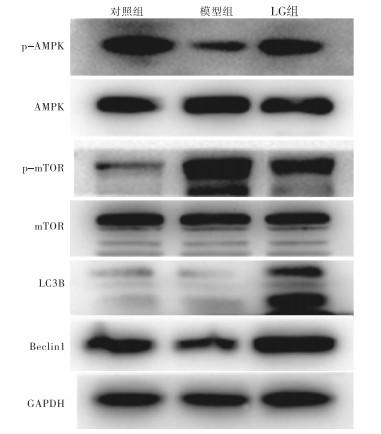
结果显示,与对照组相比,模型组p-AMPK、LC3B、Beclin1的蛋白表达下降,p-mTOR表达升高;而LG组与模型组相比,p-AMPK、LC3B、Beclin1的蛋白表达水平均升高,p-mTOR表达下降,差异均有统计学意义(P < 0.05~P < 0.01)(见图 3、表 2)。

图 3 Western blotting法检测各组肝细胞内AMPK/mTOR、LC3B、Baclin1蛋白表达
分组 n p-AMPK/AMPK p-mTOR/TOR LC3B Beclin1 对照组 10 1.10±0.14 0.48±0.09 0.37±0.10 0.79±0.20 模型组 10 0.54±0.08** 0.98±0.09** 0.26±0.03* 0.64±0.09* LG组 10 0.79±0.14**## 0.82±0.12**## 1.14±0.17**## 1.25±0.12**## F — 53.48 61.17 175.35 48.84 P — < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 0.015 0.010 0.013 0.020 q检验:与对照组相比*P < 0.05,**P < 0.01;与模型组相比##P < 0.01 表 2 各组小鼠肝细胞内p-AMPK、p-mTOR、LC3B、Baclin1蛋白水平的比较(x±s)
-
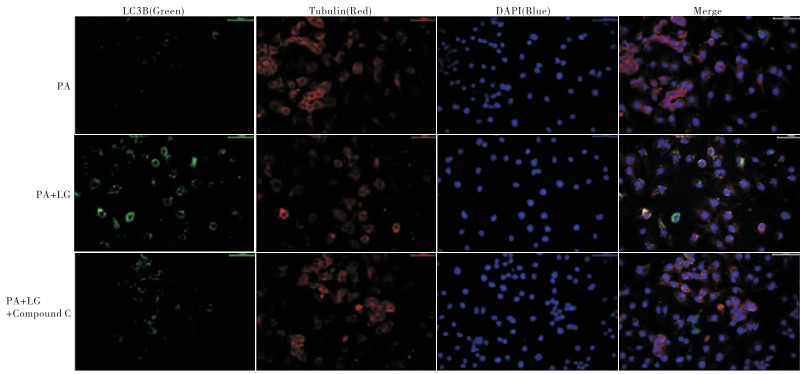
结果显示,模型组HepG2细胞可见极少量的LC3B的表达,LG组细胞内LC3B的表达较模型组升高,而AMPK抑制剂组LC3B的表达较LG组减少(见图 4)。

图 4 免疫荧光法观察各组HepG2细胞LC3B的表达
-
结果显示,和模型组相比,LG组p-AMPK、LC3B、Beclin1的蛋白表达升高,而应用AMPK抑制剂Compound C后上述指标表达较LG组受抑制下降(P < 0.05~P < 0.01);此外,和模型组相比,p-mTOR在LG组表达降低,应用AMPK抑制剂Compound C后表达较LG组增加,差异均有统计学意义(P < 0.05~P < 0.01)(见图 5、表 3)。

图 5 Western blotting检测各组HepG2细胞AMPK通路和自噬相关蛋白表达
分组 p-AMPK/AMPK p-mTOR/ mTOR LC3B Beclin1 模型组 0.70±0.05 0.96±0.12 0.70±0.03 0.52±0.03 LG组 1.01±0.07* 0.52±0.05** 1.03±0.11** 1.06±0.10** AMPK抑制剂组 0.79±0.17# 0.68±0.04**# 0.70±0.05## 0.79±0.03**## F 6.39 24.39 20.56 57.39 P < 0.05 < 0.01 < 0.01 < 0.01 MS组内 0.012 0.006 0.005 0.004 q检验:与模型组相比*P < 0.05,**P < 0.01;与LG组相比#P < 0.05,##P < 0.01 表 3 各组HepG2细胞p-AMPK、p-mTOR、LC3B、Baclin1蛋白水平的比较(x±s)
-
一项针对中国人群的分析数据显示[8],我国NAFLD的患病率达29.2%。NAFLD为糖尿病、心血管疾病的高危因素[9-10],T2DM病人中49%~62%的人群合并有NAFLD,肥胖T2DM病人中NAFLD的患病率高达70%。NAFLD主要的病理演变过程为肝脏脂肪变性、肝细胞炎性改变严重发生肝细胞坏死。因此,延缓NAFLD进展,改善肝细胞脂肪变性,受到了人们的广泛重视。
LG作为长效GLP-1类似物,不仅具有GLP-1降脂、减重、降压等作用[11-12],而且在保留其生物活性的情况下延长了半衰期[13],在2型糖尿病和心血管疾病的防治上有明确的优势。有研究[14]表明GLP-1受体激动剂可改善肝细胞内脂质沉积,发挥降脂的作用。HAO等[15]研究发现,HFD诱导的脂肪性肝炎(NASH)小鼠体质量增加,肝脂肪水平增加。LG治疗后可显著降低HFD喂养小鼠的体质量,改善肝脏脂质蓄积,抑制血清总胆固醇和LDL水平的升高。我们的研究显示,应用HFD喂养构建立肝脂肪变性的小鼠模型,发现HFD喂养的小鼠血脂升高、肝功能明显损坏,而LG能改善这一情况。HE染色和油红染色进一步观察发现肝脂肪变性的小鼠肝细胞排列紊乱、细胞膨胀、可见大量脂肪空泡形成和炎性浸润,肝组织内出现大量脂滴,而应用LG后脂肪空泡、脂滴明显减少。以上说明,LG可以改善高脂诱导的肝细胞脂肪变性,抑制肝细胞脂质沉积。
自噬是细胞自我更新的过程,分为选择性自噬和非选择性自噬。选择性自噬是细胞特异性的吞噬低能的细胞器,例如线粒体受损,细胞内脂质过量。LC3B作为自噬小体形成过程中的招募蛋白存在于自噬小体的膜上,可以追踪自噬的形成;Beclin1是哺乳动物自噬过程的特异基因,不参与自噬泡形成,是一种自噬相关调节蛋白。自噬激活后自噬体形成,可与溶酶体融合后释放的酸性脂肪酶将脂质进行降解。自噬与脂质代谢、胰岛素抵抗的相关性在很多研究中报道,自噬可以促进肝细胞中脂类物质的降解, 减少其在肝脏的集聚,最终改善脂肪变性[16]。ZHANG等[17]通过小鼠体内实验证明,通过营养缺乏诱导自噬激活,刺激LC3B与脂滴的结合,介导肝细胞内脂滴的降解,自噬增强可以保护肝细胞。长期HFD导致体内游离脂肪酸(FFA)增加,肝细胞自噬减弱,脂质蓄积最终形成脂肪肝。近期研究显示,LG可以改善FFA诱导的脂毒性肝细胞损伤,并通过促进细胞自噬来改善NAFLD中肝细胞脂肪变性[18]。LG可以诱导HFD小鼠自噬,也可以通过增强自噬通量减轻PA诱导的脂质积聚[19]。本研究无论动物实验还是细胞实验结果均显示LG应用后与模型组比较,自噬蛋白LC3B、Beclin1表达均明显增多,初步探讨了自噬在LG改善肝细胞脂肪变性的作用。
mTOR是自噬主要抑制通路之一,是AMPK下游信号分子。AMPK的激活会使Beclin1发生磷酸化,促进自噬的发生和发展。AMPK是自噬的经典上游调节剂,能被GLP-1R激动剂激活。有研究[20]显示AMPK是通过抑制肝脏细胞新生脂肪产生、增加肝脏脂肪酸的氧化能力、保护肝脏脂肪组织中线粒体功能这3种方式改善NAFLD肝脏脂肪变性。AMPK/mTOR介导的自噬水平在高脂喂养的小鼠及游离脂肪酸处理的LO2细胞(人正常肝细胞)中均有明显抑制作用[21]。小鼠HFD16周后肝脏自噬水平下调,机制考虑与p-AMPK/mTOR信号通路活性抑制相关[22]。以上动物及细胞实验均提示AMPK/mTOR可以诱导自噬改善肝脏细胞脂肪变性,我们的实验Western blotting结果中发现,自噬标志物LC3B等蛋白表达增加,AMPK/mTOR信号通路活化,两者之间呈现一致性改变。应用免疫荧光染色和Western blotting发现模型组自噬的表达很低,LG组明显增高,而AMPK抑制剂Compound C能拮抗LG对自噬的增强作用。最终我们得出结论,肝细胞变性时应用LG所产生的自噬增加与AMPK/mTOR信号通路存在相关性。LG能通过活化AMPK抑制mTOR促进自噬的发生,改善肝细胞脂肪变性。AMPK抑制剂能拮抗这一反应的作用。我们的研究结果有助于为临床应用LG治疗NAFLD及其相关代谢综合征提供证据。
利拉鲁肽通过AMPK/mTOR信号通路诱导自噬改善肝脂肪变性
Liraglutide induces autophagy through AMPK / mTOR signaling pathway and improves hepatocyte steatosis
-
摘要:
目的探讨腺苷酸活化蛋白激酶(AMPK)/雷帕霉素靶蛋白(mTOR)通路在利拉鲁肽(LG)改善肝细胞脂肪变性中的作用及机制。 方法动物实验:将小鼠分为对照组、模型组和LG组,各10只,对照组普通饲料喂养, 另2组高脂饲料喂养建立肝脂肪变性模型。LG组应用LG腹腔注射0.6 mg·kg-1·d-1,另2组应用等量0.9%氯化钠溶液,连续4周。检测小鼠血清丙氨酸氨基转移酶(ALT)、天冬氨酸氨基转移酶(AST)、低密度脂蛋白(LDL)、胆固醇(TC)、三酰甘油(TG);应用HE染色和油红O染色观察肝细胞形态、脂肪变性和脂滴情况;应用蛋白质印迹(Western blotting)法检测p-AMPK、p-mTOR及自噬蛋白LC3B、Beclin1表达情况。细胞实验:应用棕榈酸(PA)诱导建立HepG2肝细胞脂肪变性模型,分为模型组(PA)、LG组(PA+LG)和AMPK抑制剂组(PA+LG+Compound C)。应用免疫荧光检测自噬相关蛋白LC3B的表达;应用Western blotting法检测p-AMPK、p-mTOR、LC3B、Beclin1的蛋白表达情况。 结果动物实验:模型组ALT、AST、LDL、TC、TG升高,应用LG后下降,差异均有统计学意义(P < 0.05~P < 0.01);HE染色、油红O染色发现,模型组肝细胞可见大片脂肪变性、大量融合脂滴,LG组脂变减轻、脂滴含量减少。Western blotting结果显示,模型组p-AMPK、LC3B、Beclin1的蛋白表达下降,而LG组中升高,模型组p-mTOR升高,而LG组中降低(P < 0.05~P < 0.01)。细胞实验:免疫荧光染色LC3B发现,LG组较模型组增加,而AMPK抑制剂组,LC3B的表达受到抑制;Western blotting结果示:和模型组相比,LG组p-AMPK、LC3B、Beclin1的蛋白表达升高,而应用AMPK抑制剂后表达受抑制,而p-mTOR相反, 差异均有统计学意义(P < 0.05~P < 0.01)。 结论LG通过AMPK/mTOR通路来改善肝细胞脂肪变性,自噬可能参与其中。 -
关键词:
- 肝脂肪变性 /
- 利拉鲁肽 /
- 腺苷酸活化蛋白激酶/雷帕霉素靶蛋白通路 /
- 自噬
Abstract:ObjectiveTo investigate the role and possible mechanism of adenylate activated protein kinase(AMPK)/rapamycin target protein(mTOR) pathway in the improvement of liraglutide(LG) on hepatocyte steatosis. MethodsIn animal experiment, the mice were randomly divided into control group, model group and LG group, with 10 mice in each group.The mice in the control group were fed with ordinary diet, and the mice in other groups were fed with high-fat diet to construct hepatic steatosis model.In the LG group, the mice were injected with LG at 0.6 mg·kg-1·d-1 intraperitoneally, and in the other two groups, the mice were treated with the same amount of 0.9% sodium chloride solution for 4 weeks.The levels of serum alanine aminotransferase(ALT), aspartate aminotransferase(AST), low density lipoprotein(LDL), cholesterol(TC) and triglyceride(TG) were detected; the changes of morphology, steatosis and lipid droplets of hepatocytes were observed through HE and oil red O staining methods.The expressions of p-AMPK, p-mTOR, autophagy-related proteins LC3B and Beclin1 were detected by Western blotting.In cell experiment, the palmitic acid(PA) induced steatosis model in HepG2 hepatocytes was established, the cells were equally divided into model group(PA), LG Group(PA+LG) and AMPK inhibitor group(PA+LG+compound C).The expression of autophagy-related protein LC3B was detected by immunofluorescence; the protein expressions of p- AMPK, p-mTOR, LC3B and Beclin1 were detected by Western blotting. ResultsIn contrast to control group, the levels of ALT, AST, LDL, TC and TG in model group were increased significantly, and decreased significantly after LG treatment (P < 0.05 to P < 0.01);HE and oil red O staining showed that in the model group, there had large areas of steatosis and a large number of fused lipid droplets.The steatosis and lipid droplets content was decreased significantly in the LG group.Western blotting results showed that the protein expressions of p-AMPK, LC3B and Beclin1 were decreased significantly in model group, but increased in LG group, p-mTOR was increased in model group, and decreased in LG group(P < 0.05 to P < 0.01).Cell experiment: immunofluorescence staining showed that LC3B expression in LG group was significantly higher than that in model group, while LC3B expression was inhibited in AMPK inhibitor group.Western blotting results showed that compared with the model group, the protein expressions of p-AMPK, LC3B and Beclin1 in LG group were significantly increased, but the expressions were inhibited after the application of AMPK inhibitor, while p-mTOR expression was on the contrary, which had statistically significance(P < 0.05 to P < 0.01). ConclusionsLiraglutide improved hepatocyte steatosis through AMPK/mTOR pathway, and autophagy may be involved. -
表 1 各组小鼠ALT、AST、LDL、TC、TG比较(x±s)
分组 n ALT/(U/L) AST/(U/L) LDL/(mmo/L) TC/(mmol/L) TG/(mmol/L) 对照组 10 11.33±1.75 31.04±4.26 0.44±0.09 2.04±0.17 0.34±0.05 模型组 10 107.80±7.33** 63.49±8.90** 1.04±0.15** 5.02±0.55** 1.22±0.18** LG组 10 40.40±3.05**## 40.27±6.02**## 0.57±0.12*## 3.57±0.45**## 0.46±0.11*## F — 1 111.98 62.79 68.54 126.41 144.35 P — < 0.01 < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 22.022 44.523 0.015 0.176 0.016 q检验:与对照组相比*P < 0.05,**P < 0.01;与模型组相比##P < 0.01  下载: 导出CSV
下载: 导出CSV
表 2 各组小鼠肝细胞内p-AMPK、p-mTOR、LC3B、Baclin1蛋白水平的比较(x±s)
分组 n p-AMPK/AMPK p-mTOR/TOR LC3B Beclin1 对照组 10 1.10±0.14 0.48±0.09 0.37±0.10 0.79±0.20 模型组 10 0.54±0.08** 0.98±0.09** 0.26±0.03* 0.64±0.09* LG组 10 0.79±0.14**## 0.82±0.12**## 1.14±0.17**## 1.25±0.12**## F — 53.48 61.17 175.35 48.84 P — < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 0.015 0.010 0.013 0.020 q检验:与对照组相比*P < 0.05,**P < 0.01;与模型组相比##P < 0.01
下载: 导出CSV
表 3 各组HepG2细胞p-AMPK、p-mTOR、LC3B、Baclin1蛋白水平的比较(x±s)
分组 p-AMPK/AMPK p-mTOR/ mTOR LC3B Beclin1 模型组 0.70±0.05 0.96±0.12 0.70±0.03 0.52±0.03 LG组 1.01±0.07* 0.52±0.05** 1.03±0.11** 1.06±0.10** AMPK抑制剂组 0.79±0.17# 0.68±0.04**# 0.70±0.05## 0.79±0.03**## F 6.39 24.39 20.56 57.39 P < 0.05 < 0.01 < 0.01 < 0.01 MS组内 0.012 0.006 0.005 0.004 q检验:与模型组相比*P < 0.05,**P < 0.01;与LG组相比#P < 0.05,##P < 0.01
下载: 导出CSV
-
[1] ARAB JP, ARRESE M, TRAUNER M. Recent insights into the pathogenesis of nonalcoholic fatty liver disease[J]. Annu Rev Pathol, 2018, 13(1): 321. doi: 10.1146/annurev-pathol-020117-043617 [2] MVLLER TD, FINAN B, BLOOM SR, et al. Glucagon-like peptide 1(GLP-1)[J]. Mol Metab, 2019, 30: 72. doi: 10.1016/j.molmet.2019.09.010 [3] LYU J, IMACHI H, FUKUNAGA K, et al. Role of ATP-binding cassette transporter A1 in suppressing lipid accumulation by glucagon-like peptide-1 agonist in hepatocytes[J]. Mol Metab, 2020, 34: 16. doi: 10.1016/j.molmet.2019.12.015 [4] ZHAO B, QIANG L, JOSEPH J, et al. Mitochondrial dysfunction activates the AMPK signaling and autophagy to promote cell survival[J]. Genes Dis, 2016, 3(1): 82. doi: 10.1016/j.gendis.2015.12.002 [5] WANG H, LIU Y, WANG D, et al. The upstream pathway of mtor-mediated autophagy in liver diseases[J]. Cells, 2019, 8(12): 1597. doi: 10.3390/cells8121597 [6] TONG W, JU L, QIU M, et al. Liraglutide ameliorates non-alcoholic fatty liver disease by enhancing mitochondrial architecture and promoting autophagy through the SIRT1/SIRT3-FOXO3a pathway[J]. Hepatol Res, 2016, 46(9): 933. doi: 10.1111/hepr.12634 [7] YAN J, YAO B, KUANG H, et al. Liraglutide, sitagliptin, and insulin glargine added to metformin: the effect on body weight and intrahepatic lipid in patients with type 2 diabetes mellitus and nonalcoholic fatty liver disease[J]. Hepatology, 2019, 69(6): 2414. doi: 10.1002/hep.30320 [8] 中华医学会内分泌学分会, 中华医学会糖尿病学分会. 中国成人2型糖尿病合并非酒精性脂肪性肝病管理专家共识[J]. 中华内分泌代谢杂志, 2021, 37(7): 589. doi: 10.3760/cma.j.cn311282-20210105-00016 [9] HAZLEHURST JM, WOODS C, MARJOT T, et al. Non-alcoholic fatty liver disease and diabetes[J]. Metabolism, 2016, 65(8): 1096. doi: 10.1016/j.metabol.2016.01.001 [10] NIEDERSEER D, WERNLY S, BACHMAYER S, et al. Diagnosis of non-alcoholic fatty liver disease (NAFLD) is independently associated with cardiovascular risk in a large austrian screening cohort[J]. J Clin Med, 2020, 9(4): 1065. doi: 10.3390/jcm9041065 [11] SU K, YI B, YAO BQ, et al. Liraglutide attenuates renal tubular ectopic lipid deposition in rats with diabetic nephropathy by inhibiting lipid synthesis and promoting lipolysis[J]. Pharmacol Res, 2020, 156: 104778. [12] RONDINELLI M, ROSSI A, GANDOLFI A, et al. Use of liraglutide in the real world and impact at 36 months on metabolic control, weight, lipid profile, blood pressure, heart rate, and renal function[J]. Clin Ther, 2017, 39(1): 159. doi: 10.1016/j.clinthera.2016.11.001 [13] RUSSELL-JONES D. Molecular, pharmacological and clinical aspects of liraglutide, a once-daily human GLP-1 analogue[J]. Mol Cell Endocrinol, 2009, 297(1): 137. [14] TAHER J, BAKER CL, CUIZON C, et al. GLP-1 receptor agonism ameliorates hepatic VLDL overproduction and de novo lipogenesis in insulin resistance[J]. Mol Metab, 2014, 3(9): 823. doi: 10.1016/j.molmet.2014.09.005 [15] HAO T, CHEN H, WU S, et al. LRG ameliorates steatohepatitis by activating the AMPK/mTOR/SREBP1 signaling pathway in C57BL/6J mice fed a highfat diet[J]. Mol Med Rep, 2019, 20(1): 701. [16] 沈敏燕, 胡曦, 朱静, 等. 黄连碱促进肝细胞自噬及胆固醇外流改善脂肪肝的脂质蓄积[J]. 蚌埠医学院学报, 2021, 46(4): 421. [17] ZHANG E, YIN S, SONG X, et al. Glycycoumarin inhibits hepatocyte lipoapoptosis through activation of autophagy and inhibition of ER stress/GSK-3-mediated mitochondrial pathway[J]. Sci Rep, 2016, 6: 38138. [18] 张强, 刘勤, 牛春燕. 利拉鲁肽减轻脂毒性肝细胞损伤并促进自噬改善非酒精性脂肪肝[J]. 中华肝脏病杂志, 2021, 29(5): 456. [19] FANG Y, JI L, ZHU C, et al. Liraglutide alleviates hepatic steatosis by activating the tfeb-regulated autophagy-lysosomal pathway[J]. Front Cell Dev Biol, 2020, 8: 602574. [20] ALTAMIRANO J, QI Q, CHOUDHRY S, et al. Non-invasive diagnosis: non-alcoholic fatty liver disease and alcoholic liver disease[J]. Transl Gastroenterol Hepatol, 2020, 5: 31. [21] ZHANG S, MAO Y, FAN X. Inhibition of ghrelin oacyltransferase attenuated lipotoxicity by inducing autophagy via AMPK-mTOR pathway[J]. Drug Design, Develop Ther, 2018, 12(4): 873. [22] 徐玲, 马红艳, 杨军, 等. 高脂饮食通过p-AMPK/mTOR信号通路下调小鼠肝细胞自噬水平[J]. 基础医学与临床, 2018, 38(1): 37. -

 点击查看大图
点击查看大图

图(5)表(3)
计量
- 文章访问数: 2750
- HTML全文浏览量: 1497
- PDF下载量: 18
- 被引次数: 0
