



-
NLRP3炎症小体是一个多聚蛋白复合物,由NLRP3、ASC和Pro-caspase-1蛋白组成,可识别病原微生物和多种危险信号[1-2]。其活化后介导炎性细胞因子,如白细胞介素(IL)-1β、IL-18的成熟与分泌,在抗感染免疫和多种人类炎性相关疾病,如痛风、2型糖尿病、阿尔茨海默病等中发挥重要作用[3-4]。目前,NLRP3炎症小体相关炎性疾病的治疗策略主要针对NLRP3炎症小体活化后的产物如IL-1β,但此方法无法治疗NLRP3炎症小体活化后IL-18分泌和细胞焦亡导致的疾病,且易影响细胞其他生理功能,存在较多弊端[5]。靶向抑制NLRP3炎症小体活化成为治疗相关炎性疾病的新策略,多种NLRP3炎症小体抑制剂表现出良好效果[6-9],但还难以应用于临床。桑黄酮是具有抗炎抗氧化中草药桑枝中的主要活性成分,已被证实在神经炎症上有明显抑制作用[10-11],但其能否抑制NLRP3炎症小体活化并缓解相关炎性疾病有待证明。本文旨在探究桑黄酮对NLRP3炎症小体活化及其相关炎性疾病的作用。
-
SPF级C57BL/6(B6)雄性小鼠购自江苏集萃药康生物科技股份有限公司。造模时体质量22~24 g,鼠龄7~10周,在无特定病原体(SPF)条件下饲养。人急性单核细胞白血病THP-1细胞系,受赠于蚌埠医学院汪洪涛教授课题组。本研究经蚌埠医学院伦理委员会审核批准(审批号:伦动科批字[2020]第057号)。
-
桑黄酮(Mul,上海陶素生化公司,货号:TQ0161)、巨噬细胞集落刺激因子(M-CSF)(中国novoprotein公司,货号:CB34)、PMA(SIGMA公司,货号:79346)、脂多糖(LPS)(Invivogen公司,货号:tlrl-eklps)、尼日利亚菌素(Nigericin,SIGMA公司,货号:N7143)、腺嘌呤核苷三磷酸(ATP)(SIGMA公司,货号:A2383)、红细胞裂解液(碧云天公司,货号:C3702)、PBS(Servicebio公司,货号:G0002)、DMEM细胞培养基(Gibco公司,货号:C11965500BT)、RPMI 1640培养基(Gibco公司,货号:C11875500BT)、ELISA试剂盒[小鼠IL-1β、IL-6、肿瘤坏死因子(TNF-α)、人IL-1β ELISA kit,购自R&D公司]、Western blotting抗体[抗小鼠Caspase-1(p20),购自AdipoGen公司;抗人Caspase-1(p20),购自CST公司;β-actin抗体,为Abmart公司产品;山羊抗兔IgG和山羊抗小鼠IgG,购自Jackson IR公司]。
-
Thermo3543 CO2培养箱(美国ThermoFisher);Western blotting电泳转膜槽(美国BIO-RAD);ChemiDoc XRS+化学发光凝胶成像仪(美国BIO-RAD);SCI100MiniH恒温金属浴(中国SCILOGEX);CH20 BIMF200倒置显微成像仪(日本OLYMPUS);MultiskanSky多功能酶标仪(美国ThermoFisher);5810R大容量低温离心机(德国eppendorf);D3024R控温离心机(中国SCILOGEX)。
-
桑黄酮25 mg,溶于DMSO,配制为50 mmol/L贮存液,分装冻存于-80 ℃冰箱,动物实验时用PBS稀释桑黄酮贮存液,按照25 mg/kg体质量的剂量,每只鼠腹腔注射0.2 mL。单钠尿酸盐(MSU)制备:每1.68 g粉末溶于500 mL的0.01 mol/L NaOH溶液中,加热至70 ℃溶解,0.22 μm滤器过滤后室温下使MSU晶体重新析出。再用无水乙醇洗晶体并高压灭菌后烘干,无菌PBS重悬晶体至50 μg/μL,并用超声仪震荡过夜,显微镜下观察MSU晶体大小并确认其可以进入细胞,分装冻存于-20 ℃冰箱。LPS:溶于无菌PBS,配置为5 mg/mL贮存液,分装冻存于-80 ℃冰箱,动物实验时用PBS稀释,按照20 mg/kg体质量的剂量,每只鼠腹腔注射0.3 mL。
-
麻醉下处死小鼠取小鼠后腿,无菌分离腿骨骨髓细胞。加入M-CSF(50 ng/mL),37 ℃细胞培养箱分化5~7 d。培养皿内贴壁细胞即为实验所需骨髓来源巨噬细胞(BMDM)。人THP-1细胞,加入PMA(400 ng/mL)培养过夜,使得原始THP-1细胞分化为可实验用巨噬细胞。
-
已分化的BMDM或THP-1细胞,转入细胞培养板中,37 ℃培养箱培养过夜。加入LPS(100 ng/mL)预刺激3 h,加入桑黄酮30 min后,分别加入不同的NLRP3炎症小体激动剂刺激相应时间(Nigericin、ATP刺激约30 min,MSU刺激约3 h),收集细胞培养上清至EP管中待用,向12孔细胞培养板内加入细胞裂解液,裂解5 min,转入EP管中待用。
-
细胞上清需利用甲醇和氯仿抽提蛋白,最后加入上样缓冲液裂解;细胞直接用上样缓冲液裂解。裂解液金属浴101 ℃煮10 min,后经过聚丙烯酰胺凝胶电泳、转膜,5% BSA封闭1 h,一抗caspase-1(p20)(1∶ 1 000稀释),4 ℃摇床过夜,PBST洗3次,相应HRP标记二抗(1∶ 10 000稀释)室温孵育1 h,PBST洗3次,加底物显色,并用化学发光成像仪曝光拍照。
-
按试剂盒说明书在96孔ELISA板中进行,PBS稀释包被抗体(1∶ 200稀释)室温过夜孵育,PBST洗3次,10%血清封闭1 h,PBST洗3次,加入标准品和样品室温孵育2 h,PBST洗3次,检测抗体(1∶ 100稀释)孵育1 h,PBST洗3次,HRP(1∶ 50稀释)孵育30 min,PBST洗5次,加入底物显色,10%硫酸终止,酶标仪在450 nm读数。根据标准曲线计算样品中细胞因子浓度。
-
SPF级C57BL/6小鼠饲养至7~10周,小鼠体质量约22~24 g,称量体质量。(1)随机分为安慰剂组(PBS组)(3只)、感染性休克组(LPS组)(6只)、桑黄酮干预组(LPS+Mul组)(6只),桑黄酮干预组提前1 h腹腔注射桑黄酮溶液0.2 mL,剂量50 mg/kg体质量,再对LPS组和LPS+Mul组同时腹腔注射LPS(20 mg/kg)溶液0.3 mL。LPS注射4 h后,眼球取血,分离血清,颈椎脱臼处死小鼠并收集灌洗液。(2)随机分为安慰剂组(PBS组)(5只)、感染性休克组(LPS组)(10只)、桑黄酮干预组(LPS+Mul组)(10只),桑黄酮干预组腹腔注射桑黄酮溶液0.2 mL(50 mg/kg),1 h后,感染性休克组和桑黄酮干预组同时腹腔注射0.3 mL LPS(20 mg/kg)。观察并记录小鼠生存时间。
-
处死小鼠,剪开小鼠腹部外皮,用1 mL胰岛素注射针注射1 mL无菌PBS入小鼠腹腔,用手指对小鼠腹部进行2 min按摩,用25 G针头配合1 mL注射器管,吸出腹腔液,离心,上清用于ELISA检测。
-
采用t检验和生存曲线分析(Log-Rank检验)。
-
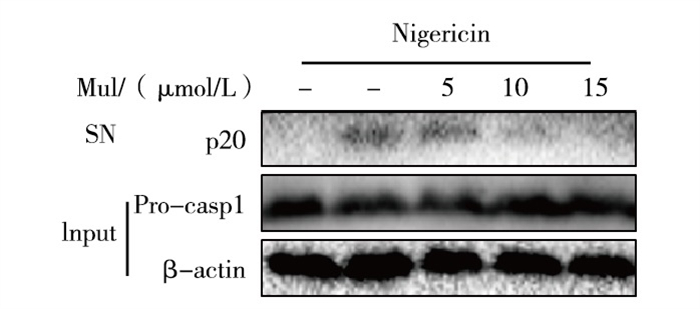
Western blotting及ELISA结果显示,桑黄酮可剂量依赖地抑制尼日利亚菌素诱导的Caspase-1(p20)表达及IL-1β的分泌(P < 0.01)(见图 1、表 1),但非炎症小体相关的细胞因子TNF-α和IL-6分泌差异无统计学意义(P>0.05)(见表 2)。

图 1 桑黄酮对Nigericin刺激的BMDM细胞p20表达的影响(SN: 细胞上清,Input: 细胞裂解液)
分组 n IL-1β t* P* mock 3 52.15±2.21 — — LPS 3 55.90±1.58 — — LPS+Nigericin 3 1 352.42±77.10 — — LPS+Nigericin+Mul(5 μmol/L) 3 921.34±38.80 8.65 < 0.01 LPS+Nigericin+Mul(10 μmol/L) 3 633.80±79.56 11.23 < 0.01 LPS+Nigericin+Mul(15 μmol/L) 3 289.15±2.67 23.87 < 0.01 *示与LPS+Nigericin组比较假设检验的结果 表 1 桑黄酮对Nigericin刺激的BMDM细胞IL-1β表达的影响(x±s;pg/mL)
分组 n TNF-α IL-6 表达水平 TNF-α t* 表达水平 TNF-α t* mock 3 79.49±7.95 — — 94.73±0.27 — — LPS 3 2 538.39±102.69 0.18 >0.05 1 065.18±90.27 0.66 >0.05 LPS+Nigericin 4 2 524.76±98.79 — — 1 025.40±70.43 — — LPS+Nigericin+Mul
(5 μmol/L)4 2 572.00±140.36 0.55 >0.05 1 016.42±50.67 0.21 >0.05 LPS+Nigericin+Mul
(10 μmol/L)4 2 409.76±176.91 1.14 >0.05 1 080.59±14.01 1.54 >0.05 LPS+Nigericin+Mul
(15 μmol/L)4 2 430.84±181.51 0.91 >0.05 1 072.95±99.90 0.78 >0.05 *示与LPS+Nigericin组比较假设检验的结果 表 2 桑黄酮对Nigericin刺激的BMDM细胞TNF-α和IL-6表达的影响(x±s;pg/mL)
-
除Nigericin外,多种病原菌和危险信号均可引起NLRP3炎症小体活化如MSU、ATP等。结果显示桑黄酮明显抑制Nigericin、ATP及MSU三种经典的NLRP3炎症小体激动剂所诱导的Caspase-1(p20)表达及IL-1β分泌(P < 0.01)(见图 2、表 3)。

图 2 桑黄酮对多种刺激剂活化的BMDM细胞p20表达的影响(SN: 细胞上清, Input : 细胞裂解液)
分组 n IL-1β t P mock
Mul(15 μmol/L)3
353.83±5.13
55.79±4.630.49 >0.05 LPS+Nigericin
LPS+Nigericin+Mul(15 μmol/L)3
31 265.48±81.23
165.96±5.9233.38* < 0.01* LPS+ATP
LPS+ATP+Mul(15 μmol/L)3
32 135.70±80.80
596.73±55.4527.20△ < 0.01△ LPS+MSU
LPS+MSU+Mul(15 μmol/L)3
31 663.46±80.84
257.73±49.1025.74# < 0.01# *示与LPS+Nigericin组比较假设检验的结果; △示与LPS+ATP组比较,#示与LPS+MSU组比较 表 3 桑黄酮对多种刺激剂活化的BMDM细胞IL-1β表达的影响(x±s;pg/mL)
-
为了明确桑黄酮在人THP-1细胞中是否同样抑制NLRP3炎症小体的活化,利用Nigericin刺激分化后的人THP-1细胞进行实验,结果显示桑黄酮可以剂量依赖性地抑制Caspase-1(p20)表达及IL-1β分泌(P < 0.01)(见图 3、表 4)。

图 3 桑黄酮对Nigericin刺激的THP-1细胞p20表达的影响(SN: 细胞上清, Input: 细胞裂解液)
分组 n IL-1β t* P* mock 3 94.73±0.27 — — LPS 3 94.58±0.47 — — LPS+Nigericin 4 1 261.93±36.16 — — LPS+Nigericin+Mul(5 μmol/L) 4 953.23±21.24 14.72 < 0.01 LPS+Nigericin+Mul(10 μmol/L) 4 586.87±25.55 30.49 < 0.01 LPS+Nigericin+Mul(15 μmol/L) 4 245.95±0.90 56.17 < 0.01 *示与LPS+Nigericin组比较假设检验的结果 表 4 桑黄酮对Nigericin刺激的THP-1细胞IL-1β表达的影响(x±s;pg/mL)
-
在小鼠感染性休克模型中进一步验证桑黄酮在动物水平上的作用,ELISA检测小鼠血清及腹腔灌洗液IL-1β及TNF-α表达水平,结果表明,与LPS组相比,LPS+Mul组小鼠血清及腹腔灌洗液中IL-1β分泌明显降低(P < 0.01),而TNF-α的水平差异无统计学意义(见表 5)。除此之外,观察LPS注射后84h内小鼠生存状态,并绘制生存曲线,结果显示LPS组小鼠14h开始死亡,36h小鼠死亡率达80%。而LPS+Mul组小鼠64h开始死亡,72h小鼠死亡率为40%(P < 0.01)(见图 4)。
分组 n 血清 腹腔灌洗液 IL-1β/(pg/mL) TNF-α/(pg/mL) IL-1β/(pg/mL) TNF-α/(pg/mL) PBS组 3 26.17±3.09 41.22±1.88 23.83±1.01 44.49±3.78 LPS组 6 275.18±69.85 190.74±14.03 527.63±65.73 85.82±13.08 LPS+Mul组 6 132.7±44.35** 177.47±10.61 288.70±88.18** 85.81±10.67 F — 23.96 191.21 52.16 16.92 P — < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 2 854.068 129.511 5 040.231 121.104 q检验:与LPS组比较** P < 0.01 表 5 桑黄酮对感染性休克小鼠血清及腹腔灌洗液IL-1β及TNF-α的影响(x±s)

图 4 桑黄酮对感染性休克小鼠生存率的影响
-
桑枝为桑科桑属植物桑的干燥嫩枝,主治风湿痹痛,水肿脚气,另有报道桑枝还具有抗炎、抗氧化及抑菌抗病毒等功效[12-14]。桑黄酮作为桑枝的主要活性成分,现有研究主要集中在神经炎症、脂质代谢等方面,有研究[15]发现其通过调节Wnt/β-catenin信号通路抑制神经炎症帕金森综合征;另有研究[16]发现桑黄酮可通过调节脂质代谢关键酶来降低肥胖的发生率。帕金森综合征和肥胖的发生发展与NLRP3炎症小体的异常活化密切相关[17-18]。目前,桑黄酮对NLRP3炎症小体作用尚不清楚,因此,探究其是否通过调节NLRP3炎症小体发挥抗炎作用对明确桑黄酮的抗炎机制具有重要意义。
NLRP3炎症小体在炎性疾病中发挥重要作用,使其成为治疗相关疾病的关键靶点。近年来,越来越多的小分子化合物被报道可以抑制NLRP3炎症小体活化,多种中草药成分被证实对抑制NLRP3炎症小体活化具有良好效果[19-21],但还难以应用于临床。寻找可靠的NLRP3炎症小体抑制剂成为新的研究方向[22]。因此,本研究主要探究中草药抗炎成分桑黄酮的抗炎作用是否通过抑制NLRP3炎症小体活化来实现。我们首先在体外探究桑黄酮是否抑制NLRP3炎症小体活化,结果显示,桑黄酮在鼠BMDM细胞中抑制Caspase-1表达及IL-1β分泌,但对primming信号介导的TNF-α和IL-6的产生没有影响,证实了桑黄酮以剂量依赖性地方式抑制Nigericin诱导的NLRP3炎症小体活化。同时,除Nigericin外,NLRP3炎症小体还可被内源性DAMPs及多种病原菌激活[23]。本研究也发现桑黄酮可以抑制其他NLRP3炎症小体刺激剂(如MSU和ATP)诱导的炎症小体活化,表明桑黄酮抑制NLRP3炎症小体具有广谱性。以上结果证实了桑黄酮可抑制鼠BMDM细胞中NLRP3炎症小体的活化。接下来笔者探讨桑黄酮在人THP-1细胞中是否抑制NLRP3炎症小体活化,结果显示,桑黄酮显著抑制Caspase-1表达及IL-1β分泌,证实了桑黄酮在人THP-1细胞中同样可以抑制NLRP3炎症小体活化。
NLRP3炎症小体的异常活化会导致大量炎性因子释放,诱导多种炎症性疾病发生,如肝纤维化[24]、炎症性肠病[25]及感染性休克[26]等。严重的感染性休克是内、外科重症监护病房中最常见的死亡原因,也是临床上亟待解决的问题之一[27]。我们构建了最常见的感染性休克模型——小鼠腹腔注射LPS,研究结果显示,桑黄酮可以明显抑制感染性休克小鼠血清和腹腔灌洗液中NLRP3炎症小体依赖的IL-1β的产生,但对NF-κB信号通路介导的TNF-α无影响。表明桑黄酮通过抑制NLRP3炎症小体活化缓解小鼠急性炎症。此外,观察LPS腹腔注射84 h内小鼠生存状态发现,LPS造模组小鼠14 h后开始死亡,而桑黄酮治疗组小鼠64 h开始死亡,桑黄酮显著延长了感染性休克小鼠的生存时间。这些结果表明桑黄酮通过抑制NLRP3炎症小体激活,在体内减轻了LPS诱导的感染性休克。
综上所述,桑黄酮不仅可以抑制体外鼠BMDM和人THP-1细胞NLRP3炎症小体活化,还可以阻抑体内NLRP3炎症小体激活,进而减轻炎症反应。明确桑黄酮对NLRP3炎症小体相关疾病的治疗作用,对开发其药用价值及推进临床应用具有重要意义。
桑黄酮抑制NLRP3炎症小体活化并缓解感染性休克的作用研究
Effect of mulberrin on inhibiting the activation of NLRP3 inflammasome and relieving infectious shock
-
摘要:
目的探究桑枝提取物桑黄酮对NLRP3炎症小体活化的影响, 为相关炎性疾病的治疗提供新思路。 方法在细胞水平上, 使用Western blotting和ELISA方法, 检测小鼠骨髓来源的巨噬细胞(BMDM)和人THP-1细胞在尼日利亚菌素、单钠尿酸盐和腺嘌呤核苷三磷酸(ATP)3种刺激剂诱导下的半胱氨酸蛋白酶-1(Caspase-1)和白细胞介素(IL)-1β表达水平。在动物实验中, 构建脂多糖(LPS)诱导的感染性休克模型, 将实验小鼠分为安慰剂组、感染性休克组和桑黄酮干预组, 利用ELISA检测小鼠腹腔灌洗液及血清中IL-1β和肿瘤坏死因子-α(TNF-α)水平。 结果在BMDM和THP-1细胞中, 桑黄酮可以抑制尼日利亚菌素、单钠尿酸盐和ATP这3种炎症小体激动剂诱导的Caspase-1和IL-1β的加工和成熟(P < 0.01), 而对非炎症小体相关的细胞因子TNF-α的分泌无影响(P>0.05)。在动物实验中, 发现桑黄酮可以缓解LPS诱导的小鼠感染性休克(P < 0.01), 明显抑制LPS诱导的小鼠血清及腹腔液中IL-1β的表达(P < 0.01), 延长LPS造模后小鼠的存活时间(P < 0.01)。 结论桑黄酮作为一种中草药活性成分提取物, 可以在体外和体内抑制NLRP3炎症小体的活化, 为NLRP3相关疾病的治疗提供了实验依据。 Abstract:ObjectiveTo investigate the effect of mulberrin, a Chinese herbal extract, on NLRP3 inflammasome activation, and provide new insight for NLRP3-related disease. MethodsWestern blotting and ELISA were used to detect the expression level of caspase-1 and interleukin-1β of mouse bone marrow derived macrophages (BMDM) and human THP-1 cells induced by three agonists: Nigerian bacteriocin, monosodium urate and adenosine triphosphate (ATP).Mouse septic shock model induced by lipopolysaccharide (LPS) was constructed, and experimental mice were divided into placebo group, septic shock group and mulberrin treatment group.The levels of IL-1β and TNF-α in intraperitoneal lavage fluid and serum were detected by ELISA. ResultsIn vitro experiments, we found that the processing and maturation of caspase-1 and IL-1β induced by three classic inflammasome agonists (Nigericin, MSU and ATP) were inhibited by mulberrin in BMDM and THP-1 cells (P < 0.01).However, there was no obviously difference observed on the secretion of non-inflammasome-related cytokines, such as TNF-α (P>0.05).In vivo experiments, mulberrin treatment alleviated septic shock, significantly inhibited IL-1β expression in serum and intraperitoneal lavage fluid of mice, and prolonged the survival time of LPS-induced mice(P < 0.01). ConclusionsAs an active ingredient of Chinese herb extract, mulberrin can inhibit the NLRP3 inflammasome activation in vitro and in vivo.Our study provide experimental evidences for the treatment of NLRP3-related diseases. -
Key words:
- mulberrin /
- NLRP3 inflammasome /
- inflammasome-related diseases
-
表 1 桑黄酮对Nigericin刺激的BMDM细胞IL-1β表达的影响(x±s;pg/mL)
分组 n IL-1β t* P* mock 3 52.15±2.21 — — LPS 3 55.90±1.58 — — LPS+Nigericin 3 1 352.42±77.10 — — LPS+Nigericin+Mul(5 μmol/L) 3 921.34±38.80 8.65 < 0.01 LPS+Nigericin+Mul(10 μmol/L) 3 633.80±79.56 11.23 < 0.01 LPS+Nigericin+Mul(15 μmol/L) 3 289.15±2.67 23.87 < 0.01 *示与LPS+Nigericin组比较假设检验的结果  下载: 导出CSV
下载: 导出CSV
表 2 桑黄酮对Nigericin刺激的BMDM细胞TNF-α和IL-6表达的影响(x±s;pg/mL)
分组 n TNF-α IL-6 表达水平 TNF-α t* 表达水平 TNF-α t* mock 3 79.49±7.95 — — 94.73±0.27 — — LPS 3 2 538.39±102.69 0.18 >0.05 1 065.18±90.27 0.66 >0.05 LPS+Nigericin 4 2 524.76±98.79 — — 1 025.40±70.43 — — LPS+Nigericin+Mul
(5 μmol/L)4 2 572.00±140.36 0.55 >0.05 1 016.42±50.67 0.21 >0.05 LPS+Nigericin+Mul
(10 μmol/L)4 2 409.76±176.91 1.14 >0.05 1 080.59±14.01 1.54 >0.05 LPS+Nigericin+Mul
(15 μmol/L)4 2 430.84±181.51 0.91 >0.05 1 072.95±99.90 0.78 >0.05 *示与LPS+Nigericin组比较假设检验的结果
下载: 导出CSV
表 3 桑黄酮对多种刺激剂活化的BMDM细胞IL-1β表达的影响(x±s;pg/mL)
分组 n IL-1β t P mock
Mul(15 μmol/L)3
353.83±5.13
55.79±4.630.49 >0.05 LPS+Nigericin
LPS+Nigericin+Mul(15 μmol/L)3
31 265.48±81.23
165.96±5.9233.38* < 0.01* LPS+ATP
LPS+ATP+Mul(15 μmol/L)3
32 135.70±80.80
596.73±55.4527.20△ < 0.01△ LPS+MSU
LPS+MSU+Mul(15 μmol/L)3
31 663.46±80.84
257.73±49.1025.74# < 0.01# *示与LPS+Nigericin组比较假设检验的结果; △示与LPS+ATP组比较,#示与LPS+MSU组比较
下载: 导出CSV
表 4 桑黄酮对Nigericin刺激的THP-1细胞IL-1β表达的影响(x±s;pg/mL)
分组 n IL-1β t* P* mock 3 94.73±0.27 — — LPS 3 94.58±0.47 — — LPS+Nigericin 4 1 261.93±36.16 — — LPS+Nigericin+Mul(5 μmol/L) 4 953.23±21.24 14.72 < 0.01 LPS+Nigericin+Mul(10 μmol/L) 4 586.87±25.55 30.49 < 0.01 LPS+Nigericin+Mul(15 μmol/L) 4 245.95±0.90 56.17 < 0.01 *示与LPS+Nigericin组比较假设检验的结果
下载: 导出CSV
表 5 桑黄酮对感染性休克小鼠血清及腹腔灌洗液IL-1β及TNF-α的影响(x±s)
分组 n 血清 腹腔灌洗液 IL-1β/(pg/mL) TNF-α/(pg/mL) IL-1β/(pg/mL) TNF-α/(pg/mL) PBS组 3 26.17±3.09 41.22±1.88 23.83±1.01 44.49±3.78 LPS组 6 275.18±69.85 190.74±14.03 527.63±65.73 85.82±13.08 LPS+Mul组 6 132.7±44.35** 177.47±10.61 288.70±88.18** 85.81±10.67 F — 23.96 191.21 52.16 16.92 P — < 0.01 < 0.01 < 0.01 < 0.01 MS组内 — 2 854.068 129.511 5 040.231 121.104 q检验:与LPS组比较** P < 0.01
下载: 导出CSV
-
[1] GONG T, LIU L, JIANG W, et al. DAMP-sensing receptors in sterile inflammation and inflammatory diseases[J]. Nat Rev Immunol, 2020, 20(2): 95. doi: 10.1038/s41577-019-0215-7 [2] FRANCHI L, MUÑOZ-PLANILLO R, NÚÑEZ G. Sensing and reacting to microbes through the inflammasomes[J]. Nat Immunol, 2012, 13(4): 325. doi: 10.1038/ni.2231 [3] GUO H, CALLAWAY JB, TING JPY. Inflammasomes: mechanism of action, role in disease, and therapeutics[J]. Nat Med, 2015, 21(7): 677. doi: 10.1038/nm.3893 [4] MARTINON F, BURNS K, TSCHOPP J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-β[J]. Mol Cell, 2002, 10(2): 417. doi: 10.1016/S1097-2765(02)00599-3 [5] DINARELLO CA, VAN DER MEER JWM. Treating inflammation by blocking interleukin-1 in humans[J]. Semin Immunol, 2013, 25(6): 469. doi: 10.1016/j.smim.2013.10.008 [6] CHEN Y, HE H, LIN B, et al. RRx-001 ameliorates inflammatory diseases by acting as a potent covalent NLRP3 inhibitor[J]. Cell Mol Immunol, 2021, 18(6): 1425. doi: 10.1038/s41423-021-00683-y [7] HE H, JIANG H, CHEN Y, et al. Oridonin is a covalent NLRP3 inhibitor with strong anti-inflammasome activity[J]. Nat Commun, 2018, 9(1): 2550. doi: 10.1038/s41467-018-04947-6 [8] HUANG Y, JIANG H, CHEN Y, et al. Tranilast directly targets NLRP3 to treat inflammasome-driven diseases[J]. EMBO Mol Med, 2018, 10(4): e8689. [9] ZHENG X, HOU Y, HE H, et al. Synthetic vitamin K analogs inhibit inflammation by targeting the NLRP3 inflammasome[J]. Cell Mol Immunol, 2021, 18(10): 2422. doi: 10.1038/s41423-020-00545-z [10] KO W, YOON CS, KIM KW, et al. Neuroprotective and anti-inflammatory effects of kuwanon C from are mediated by heme oxygenase-1 in HT22 hippocampal cells, RAW264.7 macrophage, and BV2 microglia[J]. Int J Mol Sci, 2020, 21(14): 4839. doi: 10.3390/ijms21144839 [11] XIA P, GAO X, DUAN L, et al. Mulberrin (Mul) reduces spinal cord injury (SCI)-induced apoptosis, inflammation and oxidative stress in rats via miroRNA-337 by targeting Nrf-2[J]. Biomed Pharmacother, 2018, 107: 1480. doi: 10.1016/j.biopha.2018.07.082 [12] SEO CS, LIM HS, JEONG SJ, et al. HPLC-PDA analysis and anti-inflammatory effects of Mori Cortex Radicis[J]. Nat Prod Commun, 2013, 8(10): 1443. [13] KIM TI, KWON EB, OH YC, et al. Mori ramulus and its major component morusin inhibit herpes simplex virus type 1 replication and the virus-induced reactive oxygen species[J]. Am J Chin Med, 2021, 49(1): 163. doi: 10.1142/S0192415X21500099 [14] YU W, CHEN H, XIANG Z, et al. Preparation of polysaccharides from Ramulus mori, and their antioxidant, anti-inflammatory and antibacterial activities[J]. Molecules, 2019, 24(5): 856. doi: 10.3390/molecules24050856 [15] CAO W, DONG Y, ZHAO W, et al. Mulberrin attenuates 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine (MPTP)-induced Parkinson's disease by promoting Wnt/β-catenin signaling pathway[J]. J Chem Neuroanat, 2019, 98: 63. doi: 10.1016/j.jchemneu.2019.04.003 [16] HOU XD, GE GB, WENG ZM, et al. Natural constituents from Cortex Mori Radicis as new pancreatic lipase inhibitors[J]. Bioorg Chem, 2018, 80: 577. doi: 10.1016/j.bioorg.2018.07.011 [17] GORDON R, ALBORNOZ EA, CHRISTIE DC, et al. Inflammasome inhibition prevents α-synuclein pathology and dopaminergic neurodegeneration in mice[J]. Sci Transl Med, 2018, 10(465): eaah4066. doi: 10.1126/scitranslmed.aah4066 [18] VANDANMAGSAR B, YOUM YH, RAVUSSIN A, et al. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance[J]. Nat Med, 2011, 17(2): 179. doi: 10.1038/nm.2279 [19] LIN Y, LUO T, WENG A, et al. Gallic acid alleviates gouty arthritis by inhibiting NLRP3 inflammasome activation and pyroptosis through enhancing Nrf2 signaling[J]. Front Immunol, 2020, 11: 580593. doi: 10.3389/fimmu.2020.580593 [20] 邓敏, 侯滨芬, 陈晓东. 大黄酚通过激活NLRP3炎症小体诱导胃癌细胞焦亡的机制研究[J]. 蚌埠医学院学报, 2021, 46(8): 994. [21] 周慧, 曲颂扬, 田立东. 雷公藤甲素对IgA肾病大鼠的肾保护作用及对NLRP3炎症小体的影响[J]. 蚌埠医学院学报, 2020, 45(12): 1593. [22] ZHANG X, XU A, LV J, et al. Development of small molecule inhibitors targeting NLRP3 inflammasome pathway for inflammatory diseases[J]. Eur J Med Chem, 2020, 185: 111822. doi: 10.1016/j.ejmech.2019.111822 [23] KELLEY N, JELTEMA D, DUAN Y, et al. The NLRP3 inflammasome: an overview of mechanisms of activation and regulation[J]. Int J Mol Sci, 2019, 20(13): 3328. doi: 10.3390/ijms20133328 [24] WREE A, EGUCHI A, MCGEOUGH MD, et al. NLRP3 inflammasome activation results in hepatocyte pyroptosis, liver inflammation, and fibrosis in mice[J]. Hepatology, 2014, 59(3): 898. doi: 10.1002/hep.26592 [25] BAUER C, DUEWELL P, MAYER C, et al. Colitis induced in mice with dextran sulfate sodium (DSS) is mediated by the NLRP3 inflammasome[J]. Gut, 2010, 59(9): 1192. doi: 10.1136/gut.2009.197822 [26] MAO K, CHEN S, CHEN M, et al. Nitric oxide suppresses NLRP3 inflammasome activation and protects against LPS-induced septic shock[J]. Cell Res, 2013, 23(2): 201. doi: 10.1038/cr.2013.6 [27] ANGUS DC, VAN DER POLL T. Severe sepsis and septic shock[J]. N Engl J Med, 2013, 369(9): 840. doi: 10.1056/NEJMra1208623 -

 点击查看大图
点击查看大图

图(4)表(5)
计量
- 文章访问数: 2251
- HTML全文浏览量: 1197
- PDF下载量: 21
- 被引次数: 0
